Популяция - молекула
Cтраница 1
Популяция молекул с циклическими перестановками, очевидно, мало пригодна для анализа последовательности оснований при помощи большинства существующих методов. [1]
Численность популяции молекул фермента в организме зависит от функционирования ферментобразующей системы, гормональной регуляции ее работы, сбалансированного набора аминокислот и процессов, поставляющих энергию для синтеза. Вторая группа - факторы, влияющие на активность фермента. К ним относятся природные активаторы и ингибиторы, концентрация ксфермента и субстрата. Особое значение имеет концентрация субстрата. Как правило, концентрация субстрата в организме такова, что фермент работает далеко не с полной нагрузкой. Определеннее количество молекул фермента остается резервным. Такая неполная загрузка фермента имеет, по-видимсму, вполне определенный физиологический смысл. При экстраординарных событиях, сопровождающихся резким увеличением концентрации субстрата, организм успевает обеспечить его быстрее метаболиро-вание путем включения в реакцию резервных молекул фермента. [2]
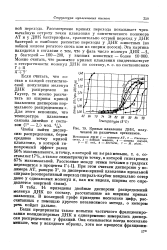 |
Кривые плавления ДНК, полученной из различных организмов. [3] |
Если считать, что состав в каждой статистической популяции молекул ДНК распределен по Гауссу, то можно рассчитать из ширины кривой плавления дисперсию нормального распределения а. Для этого вспомним, что температура плавления связана линейно с составом ( 1 - 2 5 мол. [4]
Существуют ли какие-нибудь вариации первичной структуры в пределах популяции молекул мРНК, кодирующих определенный белок. Не обнаружены у РНК бактериофагов, но описаны для одного из вирусов растений. [5]
Из данной схемы видно, что концевые повторяющиеся последовательности различаются по длине, поэтому методы определения последовательности, зависящие от гомогенности популяции молекул, применять нельзя. [6]
Форма полос преципитации позволяет судить о гомогенности или гетерогенности исследуемых белков. Гетерогенная популяция молекул обычно дает вытянутую асимметричную полосу преципитации. [7]
Если к препарате ДНК распределение молекул по составу около среднего значения не соответствует нормальному, то это отражается на форме кривой плавления. В тех случаях, когда в образцах имеются две популяции молекул с явно различными распределениями, на кривых плавления появляются два перехода ( фиг. [8]
Направленная молекулярная эволюция подобна искусственному отбору. Если надо создать молекулу, обладающую каким-либо химическим свойством, следует выбирать из большой популяции молекул те, которые в наилучшей степени выражают это свойство, и произвести из них дочерние, в разной степени похожие на родителей. [9]
При этом не предпринималось Гникаких усилий по отбору для иммобилизации каких-либо определенных участков ДНК; белки сами находили в гетерогенной популяции молекул ДНК участки своего биоспецифического связывания. [10]
В принципе точно такая же ситуация имеет место и в случае амфо-литов, например белков. Существенно отметить, во-первых, что как при рН - pi, так и при любых других аначешшх рН в огромной популяции молекул амфолитов имеются как в целом нейтральные, так и суммарно положительно и отрицательно заряженные молекулы ( с изменением рН изменяются лишь их пропорции), а, во-вторых, каков бы ни был суммарный электрический заряд молекулы амфолита ( например, белковой глобулы), на ее поверхности почти наверняка найдутся заряженные группы обоих знаков. Но здесь ее последствия могут быть более серьезными. При электрофорезе направление миграции белковой молекулы в электрическом поле определяется знаком ее суммарного заряда, а в ионообменной хроматографии на первый план выступают локальные электростатические взаимодействия. И можно себе представить такой случай, когда, например, суммарно заряженные положительно макромолекулы будут задерживаться на положительно же заряженном анионообменнике за счет взаимодействия с матрицей своих отрицательно заряженных групп, если последние, даже будучи в меньшинстве, окажутся в непосредственном соседстве с ионизированными группами обмепника, что зависит от пространственной ориентации макромолекул. [11]
Иммуноглобулины разных классов имеют много общих антигенных детерминант, но различаются по электрофоретической подвижности, поэтому они образуют при иммуноэлектрофорезе вытянутую асимметричную полосу преципитации. Особое значение этому явлению придается при диагностике так называемых моно-клональных иммуноглобулинопатий. При этих заболеваниях ( например, множественном миеломатозе или макроглобулинемии Вальденштрема) клон злокачественно пролиферирующих плазматических клеток обычно продуцирует гомогенный монокло-нальный иммуноглобулин. Патологическое увеличение популяции молекул этого иммуноглобулина приводит к тому, что при иммуноэлектрофорезе можно наблюдать характерную симметричную полосу преципитации. IgA - и IgG-типа, можно заметить также, что при значительном увеличении концентрации отдельных белков при моноклинальных иммуноглобулинопатиях происходит растворение полос преципитации в избытке антигена. Это явление может стать причиной неверного толкования иммуно-электрофореграмм, поскольку растворение преципитата воспринимается как отсутствие данной белковой фракции, хотя на самом деле она присутствует в большом избытке. В этих случаях следует устранять избыток антигена 5 - 10-кратным разбавлением исследуемой сыворотки с высокой концентрацией белка. [12]
В яоследнее время для у-глобулинов получены отпечатки пальцев после триптического переваривания. Это означает, что многие пептиды совпадают попарно и дают наложение пятен, что вполне естественно, если структурные единицы Портера А и В совпадают по структуре. В гидроли-зате популяции молекул у-глобулина наблюдается некоторое разнообразие в картине отпечатков пальцев, свидетельствующее о том, что ( - глобулины одного организма имеют разнородную первичную структуру и являются смесью химически различных, хотя и похожих, белков. [13]
В этом случае молекулы Asp заряжены в целом отрицательно; они практически не будут задерживаться на максимально отрицательно заряженном обменнике и выйдут в свободном объеме колонки. Положительный заряд Lys в среднем равен - - J-1 и в несколько раз выше, чем у Gly. Это означает, что действительно положительно заряженные молекулы будут явно преобладать в популяции молекул Lys. [14]
Возможность периодических явлений в химических реакциях неоднократно обсуждалась и в термодинамическом аспекте, причем Пригожий и Балеску [19] показали, что в условиях, когда химическая система значительно удалена от равновесия, она может совершать колебания около стационарного состояния бесконечно долго. Эти колебания Гастингс и другие считают динамической основой временной структуры клетки. Гудвин полагает, что колебания в регуляторных механизмах очень распространены, а истинные стационарные состояния составляют редкое исключение; почти все популяции молекул в живой клетке, по Гудвину, должны испытывать незатухающие колебания того или иного вида. Ссылаясь на исследования Гончарова, Рубина и Ситгривса, он приходит к выводу о малой вероятности наступления стационарных состояний в столь сложной системе, какой является клетка, и делает совершенно правильный, с нашей точки зрения, вывод, что все виды молекул в клетке находятся в динамическом состоянии - распадаются и вновь синтезируются, чем и обеспечивается поддержание незатухающих колебаний их концентраций. [15]
Страницы: 1 2