Вторичный посредник
Cтраница 2
Кроме того, пролактин влияет на секреторную активность желтого тела, а также на эритропоэз. У пресмыкающихся этот гормон контролирует рост органов и их регенерацию, у рыб - стимулирует функции щитовидной железы и семенных пузырьков. Молекулярные механизмы действия пролактина проявляются через вторичные посредники и заключаются в фосфорилировании и регуляции активности соответствующих ферментов. В медицинской практике применяют препарат л актин. Его получают из передней доли гипофиза крупного рогатого скота. Препарат усиливает лактацию в период кормления ребенка. [16]
К настоящему времени открыто более сотни различных веществ, наделенных гормональной активностью, синтезирующихся в железах внутренней секреции и регулирующих процессы обмена веществ. Установлены специфические особенности биологического действия гормонов: а) гормоны проявляют свое биологическое действие в ничтожно малых концентрациях ( от 10 - б до 10 - 12 М); б) гормональный эффект реализуется через белковые рецепторы и внутриклеточные вторичные посредники ( мессендже-ры); в) не являясь ни ферментами, ни коферментами, гормоны в то же время осуществляют свое действие путем увеличения скорости синтеза ферментов de novo или изменения скорости ферментативного катализа; г) действие гормонов в целостном организме определяется в известной степени контролирующим влиянием ЦНС; д) железы внутренней секреции и продуцируемые ими гормоны составляют единую систему, тесно связанную при помощи механизмов прямой и обратной связей. [17]
Глюкагон является гормоном-антагонистом инсулина. Он стимулирует гликогенолиз и липолиз, а также активирует процесс глюконеогенеза. Глюкагон взаимодействует с клетками-мишенями по мембрано-опосредованному механизму ( гл. Через вторичный посредник - цАМФ он активирует протеинкиназу, киназу фосфорилазу и фосфорилазу Ь, что приводит к мобилизации глюкозы из гликогена. Как и инсулин, глюкагон регулирует метаболические процессы преимущественно в печени, мышцах и жировой ткани. [18]
 |
Структура аллоксана - пиримиди-нового производного, вызывающего экспериментальный диабет у животных путем разрушения островковых клеток. [19] |
Эти рецепторы были выделены из клеточных мембран и очищены. Выделенный рецептор инсулина является специфическим гликопротеи-ном, который очень прочно связывает инсулин. Число рецепторов инсулина на поверхности клетки меняется в зависимости от условий обмена веществ; показано также, что они обладают высокой скоростью оборота. Несмотря на интенсивные исследования, проводящиеся на протяжении многих лет, до сих пор не удается обнаружить вторичный посредник, высвобождающийся при связывании инсулина с его рецепторами. [20]

Поистине, не только из ребра Адама была создана Ева. Специфические внутриклеточные рецепторы р-эстрадиола содержатся в первичных тканях-мишенях - матке и молочных железах. Эстрогенный рецептор, называемый эстрофилином I, имеет мол. При связывании молекулы эстрогена эстрофилин I претерпевает молекулярные изменения, превращаясь в эстрофилин II, который можно рассматривать как вторичный посредник в действии эстрогена. [22]
 |
Биологическая активность кортикостероидов ( Bagavan N. V., 1978. [23] |
Образование кортикостероидов осуществляется в несколько стадий, причем общим предшественником их является холестерин ( гл. Холестерин синтезируется в надпочечниках или же поступает в них из кровяного русла. В цитоплазме клеток происходит этерификация холестерина и его депонирование. Сигнал на синтез кортикостероидов формируется в гипоталамусе и реализуется в синтезе кортиколиберина. Последний, взаимодействуя с мембранными рецепторами клеток надпочечников, через систему вторичных посредников активирует эстеразу холестерола; при этом освободившийся холестерол транспортируется в митохондрии. [24]
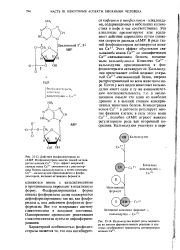
Этот эффект обусловлен связыванием ионов Са2 со специфическим Са2 - связывающим белком, называемым калъмодулином. Комплекс Са2 1 -кальмодулин присоединяется к фос-фодиэстеразе и активирует ее. Кальмодулин представляет собой недавно открытый Са2 - связывающий белок, широко распространенный во всем животном мире. Почти у всех видов животных кальмодулин имеет одну и ту же аминокислотную последовательность, т.е. в эволюционном смысле это один из наиболее древних и в высшей степени консервативных животных белков. Концентрация ионов Са2 в цитозоле регулирует многие функции клетки; в силу этого ионы Са2, подобно сАМР, играют важную регуляторную роль как вторичный посредник. [26]
Катехоламины действуют на клетки-мишени по мембрано-опосредованному механизму, чему в немалой степени способствует гидроксилирование кольца и боковой цепи этих соединений. Катехоламины взаимодействуют с а - и р-адренергическими рецепторами, локализованными в мембранах клеток-мишеней. Адреналин взаимодействует с обоими типами рецепторов, а норадреналин преимущественно с а-рецепторами. Группа ах -, а2 - рецепторов проявляет эффекты сосудосуживающего действия, сокращения гладких мышц, ингибирования липолиза. Действие ( 3-рецепторов связано с активацией аденилатциклазы, образованием цАМФ и последующим фосфорилированием белков. Например, адреналин, взаимодействуя с р-рецепторами через систему вторичных посредников, активирует протеинкиназу, которая фосфорилирует ряд цитоплазматических белков. Таким образом, адреналин регулирует гликогенолиз в печени и в мышцах, а также глюконеогенез в печени. [27]
В последние годы в связи с интенсивным изучением биологического действия нейропептидов па организм внимание исследователей было привлечено к изучению проницаемости ГЭБ для этого класса соединений. Выраженный физиологический эффект нейропептидов позволил предположить, что они легко проникают через ГЭБ. Однако многочисленные исследования свидетельствуют о крайне малом поступлении их в ЦНС; более того, рядом авторов сделано заключение о непроницаемости ГЭБ для многих пептидов. Такое, казалось бы, противоречие может быть объяснено возможностью поступления из крови и накоплением пептидов в участках мозга, но защищенных ГЭБ ( эпифиз, гипофиз, срединное возвышение, субфор-никальный орган и др.), откуда возможна диффузия в прилежащие структуры ЦНС. Из области безбарьерных зон пептиды могут проникать в ликвор мозговых желудочков или непосредственно контактировать с нервными элементами и рецепторами. К настоящему времени действительно установлено участие циклических иуклео-тидов ( вторичных посредников) в механизме действия энкефалинов. [28]
В наше время трудно назвать область естествознания, которую не интересовала бы проблема структуры и функций нуклеиновых кислот. Несмотря на огромный прогресс, достигнутый в последние десятилетия при изучении химического состава и строения нуклеиновых кислот, много проблем предстоит еще решить для выяснения зависимости между структурой и биологической ролью нуклеиновых кислот. Нет сомнения, что именно на этом пути научного поиска исследования нуклеиновых кислот будут сделаны открытия, имеющие огромное значение для биологии, медицины и всей науки о живом. Эпохальное открытие принципа комп-лементарности нуклеиновых кислот позволило проникнуть в тайны не только тонкой структуры этих биополимеров, но и механизмов синтеза и воспроизведения биологических макромолекул. Нуклеиновые кислоты выполняют ряд важных биологических функций, не свойственных другим полимерным веществам. В частности, они обеспечивают хранение и передачу наследственной информации и принимают непосредственное участие в механизмах реализации этой информации путем программирования синтеза всех клеточных белков. Структурные компоненты нуклеиновых кислот выполняют, кроме того, функции кофакторов ( коэнзим А, уридин-дифосфатглюкоза и др.), аллостерических эффекторов, входят в состав коферментов ( никотинамидадениндинуклеотид, флавинадениндинуклеотид и др.), принимая тем самым непосредственное участие в обмене веществ, а также в аккумулировании ( накоплении), переносе и трасформации энергии. Они являются предшественниками вторичных посредников ( мессенд-жеров) - циклических мононуклеотидов ( цАМФ и цГМФ), выполняющих важную функцию в передаче внутриклеточных сигналов. [29]
Страницы: 1 2