Природа - боковая цепь
Cтраница 2
Будет ли глобулярная белковая молекула раскручиваться и распластываться между поверхностью липидного слоя и водой, зависит в значительной степени от расположения и природы неполярных боковых цепей аминокислот, входящих в состав белковой молекулы, и от величины поверхностного натяжения в липид-ном слое. При малой величине поверхностного натяжения многие неполярные боковые цепи белковой молекулы проникнут в липо-фильную область двойного липидного слоя. [16]
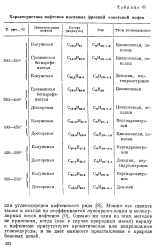 |
Характеристика нафтенов масляных фракций советской нефти. [17] |
Однако ни один из этих методов не приложим, когда ( как в случае природных масел) наряду с нафтенами присутствуют ароматические или непредельные углеводороды, и не дает никакого представления о природе боковых цепей. [18]
Этот метод, несомненно, будет весьма полезным для определения количества спиральной части в полипептидах и белках, однако еще нет полной уверенности в том, сохранится ли величина, полученная для поли - Ь - глутаминовой кислоты, при изменении природы боковых цепей. [19]
К - длина волны; Я0 - длина волны, совпадающая с длиной волны поглощения белка в УФ-области; [ а ] - величина удельного оптического вращения; ас - постоянная, характеризующая взаимодействие между молекулами растворителя и макромолекулами белка, которая учитывает содержание спиральных форм в структуре и зависит от природы боковых цепей аминокислотных веществ; Ь0 - постоянная, характеризующая Конформацию полипептидной цепи и количество спиральных участков в ней. [20]
 |
Газофазный секренатор АП-03 ( НТО АН СССР, г. Ленинград.| Схема реакционной камеры газофазного секвенаторе. [21] |
Общим требованием к субстрату для всех карбоксипептидаз является наличие а-карбоксильной группы у С-концевой аминокислоты. Природа боковой цепи у отщепляемого аминокислотного остатка - основной фактор, определяющий скорость гидролиза пептидной связи. На скорость отщепления С-концевой аминокислоты в большой степени влияет также природа соседнего с ней остатка. Расположенные в соседнем положении аминокислотные остатки с ароматической или алифатической боковой цепью, а также остатки дикарбоновых аминокислот значительно ускоряют отщепление С-концевой аминокислоты. Напротив, если в соседнем положении находятся глицин и пролин, скорость гидролиза снижается. [22]
Специфичность реакции определяется природой боковой цепи аминокислотного остатка, пространственным строением модифицируемого соединения и условиями реакции. [23]
С другой стороны, кривая зависимости давление - площадь для поливинилацетата не зависит от природы поверхности раздела и имеет одинаковый вид для поверхностей вода - воздух и вода - масло. Это указывает на большое значение природы боковых цепей для процессов расширения и сжатия монослоев, как это уже было установлено при исследовании пленок белков. Хотя поливинилацетат образует пленку растянутого типа, пленки его-сополимеров с винилстеаратом, даже если последний содержится в них в количестве всего 10 %, в значительной степени конденсированы. Это является подтверждением того, что, как упоминалось выше, конденсация пленки обусловлена когезией между неполярными группами. [24]
С другой стороны, кривая зависимости давление - площадь для поливинилацетата не зависит от природы поверхности раздела и имеет одинаковый вид для поверхностей вода - воздух и вода - - масло. Это указывает на большое значение природы боковых цепей для процессов расширения и сжатия монослоев, как это уже было установлено при исследовании пленок белков. Хотя поливинилацетат образует пленку растянутого типа, пленки его сополимеров с винилстеаратом, даже если последний содержится в них в количестве всего 10 %, в значительной степени конденсированы. Это является подтверждением того, что, как упоминалось выше, конденсация пленки обусловлена когезией между неполярными группами. [25]
Поскольку почти любое изменение молекулы изменяет электро-фильные свойства какой-нибудь ее части, метаболические изменения ФОС всегда должны сопровождаться либо активированием, либо исчезновением эффекта. К тому же, как показывает приложение 2, природа боковой цепи, соединенной с токсофорной группировкой [ такой, например, как ( RO) 2P ( S) S - ], не имеет существенного значения, если только ядовитое начало, образующееся в организме чувствительных животных, обладает мощным антихо-линэстеразным действием. Связанные с этим факторы обсуждены на стр. Поэтому можно быть достаточно свободным в выборе боковых цепей. Если известна реакция, которая не является высоко специфичной и протекает с различной интенсивностью у разных видов животных, то эти различия могут быть использованы для получения избирательной токсичности. [26]
 |
Схема переноса электронов в дыхательной цепи митохондрий. [27] |
В митохондриях обнаружено пять цитохромов ( Ь, с, с, а, а3), различающихся между собой спектрами поглощения и окислительно-восстановительными потенциалами. Различия по этим параметрам обусловлены белковыми компонентами цитохромов, природой боковых цепей их порфиринов и способом присоединения гема к белкам. Конечные цитохромы ( а а3) передают электроны на молекулярный кислород, представляя собой собственно цитохромоксидазу, в реакционном центре которой содержатся помимо двух гемов два атома меди. Образование воды имеет место при переносе на молекулу кислорода 4 электронов. Некоторые цитохромоксидазы осуществляют перенос на 02 только 2 электронов, следствием чего является возникновение перекиси водорода. Перекись водорода далее разрушается каталазой или перок-сидазой. [28]
 |
Спираль с тремя остатками на виток ( по Хагеймайеру. [29] |
Принципиально а-спираль может быть образована как L -, так и D-аминокислотнымн остатками. Склонность полипептидной цепи к образованию спиральной структуры в значительной степени зависит от природы боковых цепей аминокислот. В случае кислых и основных аминокислот дестабилизирующее действие определяется наличием зарядов на боковых группах. [30]
Страницы: 1 2 3