Проводимость - мембрана
Cтраница 2
Амплитуда ВПСП определяется двумя факторами: достигнутым уровнем МП и величиной повышения проводимости мембраны в отношении входящих по электрохимическому градиенту ионов. В латентном пейсмекерном нейроне отдельный подпороговый ВПСП вызывает дополнительно деполяризацию, составляющую волну пей-смекерного потенциала, в результате чего общий эффект деполяризации возрастает. Отличие ВПСП от наслаивающейся на него пейсмекерной волны состоит в том, что ВПСП связан с изменением сопротивления мембраны. Разделение ВПСП и наслаивающегося на него пейсмекер-ного потенциала достигается путем внутриклеточной гиперполяризации. При этом амплитуда ВПСП возрастает, а пейсмекерная волна полностью подавляется. [16]
Прямое доказательство наличия каналов состоит в том, что в отсутствие проникающих ионов Na и К высокочастотная проводимость мембраны равна нулю. Если бы в ней функционировали подвижные заряженные группы, то должен был бы наблюдаться электрический ток. Удается оценить количественные характеристики каналов, число которых, приходящееся на 1 мкм2 мембраны, составляет несколько сот. Сечение Ыа - канала - примерно 3 X 5 А2, пропускная способность-107 ион / с. [17]
В работах [53, 54] в сущности рассматривается лишь импульс / ь Кроме того, учитывается утечка тока через пассивную проводимость мембраны, дающая вклад у / гт, где гга-сопротивление мембраны единицы длины аксона в состоянии покоя. [18]
Мембрану Р - - селективного электрода изготавливают из монокристалла трифторида лантана с добавкой европия [2], которая увеличивает проводимость мембраны. В этой структуре F подвижны. В орторомбической структуре, которая не годится для электрода, каждый ион металла окружен восемью ионами. [19]
При увеличении напряжения процесс переноса ионов в большей степени лимитируется поверхностными неперемешивающимися слоями, чем объемом мембраны, поскольку проводимость мембраны резко возрастает в этих условиях. [20]
Поскольку концентрации ионов в мембране зависят от их концентраций во внешнем растворе ( или растворах), с которым мембрана находится в соприкосновении, проводимость мембран возрастает с увеличением концентрации внешнего раствора и наоборот. Таким образом, можно ожидать, что проводимость мембраны имеет низший предел, который служит характеристикой данной мембраны при малых концентрациях внешнего раствора. [21]
Существенной особенностью реакции замедления, отличающей ее от обычного тормозного действия медиатора на хемочувствительную мембрану, является то, что МП нейрона остается постоянным, а проводимость мембраны не меняется. Возникает предположение, что АХ может непосредственно влиять на механизм, ответственный за генерацию пейсмекерных потенциалов, подавляя колебательный процесс. Сходство в динамике развития и протекания реакций замедления и ускорения пейсмекерных потенциалов, как при действии тактильных раздражений, так и при прямой апликации АХ, позволяет заключить, что обе реакции, возникающие без сдвига МП, обязаны своим прохождением нейрогуморальному действию медиатора на хемочувствительную мембрану сомы нейрона вблизи пейсмекерного локуса. В этом случае АХ действует в качестве активатора-инактиватора пейсмекерного механизма. Таким образом, один и тот же медиатор в разных нейронах действует на один и тот же пейсме-керный механизм противоположным образом. Объяснение этому следует искать в том, что АХ, реагируя с разными рецепторами мембраны, образует различные по своему действию продукты реакции. Именно эти комплексы, а не сам АХ, воздействуют на механизм генерации пейсмекерных потенциалов. С этой точки зрения становится понятной роль высокой чувствительности мембраны сомы нейрона к специфическому медиатору. Возможность запуска пейсмекерной активности в латентном пейсмекерном нейроне при однократном ортодромном раздражении или однократной апликации АХ на сому нейрона показывает, сколь эффективны могут быть факторы, управляющие состоянием пейсмекерного механизма. [22]
Не исключено, что ингибиро-вание Mg 2, Na, К - АТФ-азы может явиться одним из механизмов, через который реализуется блокирующее действие МАП на проводимость нервных мембран. Известно, что Mg 2, Na, К - АТФ-аза участвует в активном транспорте ионов К и Na через биомсмбраны. [23]
Переходя к комплексным реакциям в виде ускорения-замедления и замедления-ускорения, которые вызываются тактильными или электрическими ортодромными раздражениями, следует подчеркнуть, что в этом случае уровень МП и проводимость мембраны также не меня -, ются. Логично предположить, что в этом случае в одном нейроне имеет место комбинация двух реакций - реакции ускорения и замедления. Это может быть вызвано действием разных медиаторов, конвергирующих на исследуемом нейроне, либо действием одного медиатора на два типа рецептивных белков, содержащихся в мембране и обладающих разной реактивностью и скоростью сенси-тизации-десенситизации. Убедительным доказательством последнего утверждения является то, что один и тот же медиатор ( АХ) может при микроапликации на мембрану одного и того же нейрона вызывать как реакцию ускорения, так и реакцию замедления. Последовательность событий, видимо, связана с разной скоростью сенситиза-ции-десенситизации. Реакцию ускорения-замедления можно представить как случай, когда исходная чувствительность хеморецепторов мембраны, вызывающих активацию пейсмекерной активности, повышена. Затем в результате десенситизации этих рецепторов или сенсити-зации рецепторов, ответственных за подавление пейсмекерной активности, реакция меняет свой знак и реакция ускорения переходит в реакцию замедления. [24]
На основе принятой модели показано, что развитие спайка на мембрана аксона начинается с того момента, когда мембранный потенциал достигает значения, при котором проводимость отрицательна и превышает по абсолютной величине проводимость мембраны. [25]
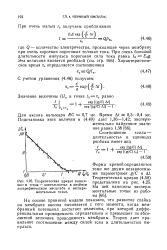 |
Теоретическая кривая зависимости сила-длительность в двойном логарифмическом масштабе и экспериментальные точки. [26] |
На основе принятой модели показано, что развитие спайка на мембране аксона начинается с того момента, когда мембранный потенциал достигает величины, при которой дифференциальная проводимость отрицательна и превышает по абсолютной величине проводимость мембраны. Теория дает правильное соотношение между силой тока и длительностью импульса. [27]
Мембрана аксона имеет сопротивление 1000 Ом - см2, ее емкость равна 1 мкФ / см2, что соответствует бимолекулярному липидному слою толщиной 50 А с диэлектрической проницаемостью е5 и удельным сопротивлением 2 - Ю9 Ом - см. Во время генерации импульса проводимость мембраны увеличивается примерно в 1000 раз. Рисунок изображает лишь один элемент мембраны, и следует представить себе длинную линейную последовательность таких элементов, моделирующих непрерывный кабель. Сопротивление R характеризует аксоплазму, наружный раствор имеется в большом избытке и изображается проводником без сопротивления. Y ответственна за движение других ионов, не изменяющееся при возбуждении. [28]
Кроме описанного действия GTX, ВТХ, вератридина, а также АТХ II на возбудимую мембрану, эти токсины стимулируют передачу химическими синапсами, что не является неожиданным, так как электрофизиологическими методами было показано, что секреция нейромедиатора ускоряется из-за деполяризации благодаря увеличению кальциевой проводимости пресинап-тической мембраны. [29]
Действие ЦП с успехом изучается на искусственных двуслойных липидных мембранах ( см. стр. Проводимость мембраны линейно возрастает с концентрацией ЦП в растворе. [30]
Страницы: 1 2 3 4