Проводимость - мембрана
Cтраница 3
 |
Схама модели переносчиков. Внизу эквивалентная схема мембраны. [31] |
Предполагается, что ионы в мембране располагаются в двух потенциальных ямах ширины К вблизи поверхностей мембраны, между которыми они совершают переходы с константами скоростей йс и их. Проводимость мембраны вычисляется на основе рассмотрения кинетики перемещения ионов. [32]
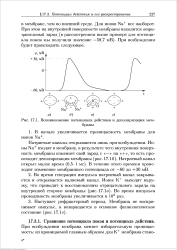 |
Возникновение потенциала действия и деполяризация мембраны. [33] |
Во время генерации импульса натриевый канал закрывается и открывается калиевый канал. Во время импульса проводимость мембраны увеличивается в 103 раз. [34]
Эти значения были получены при определении проводимости соответствующей мембраны после приведения ее в состояние равновесия с растворами различной концентрации. При низких концентрациях проводимость мембраны почти не зависит от величины концентрации раствора и равна проводимости, определяемой в то время, когда мембрана находится в равновесии с дистиллированной водой. По мере того как концентрация раствора увеличивается, проводимость мембраны, находящейся в равновесии с раствором, также увеличивается. В результате этого проводимость мембраны увеличивается. Очень важно, что в этих условиях ток уже не проводится исключительно ионами натрия, а мигрирующие ионы хлора также принимают участие в переносе тока, и мембрана не является больше чисто катионяым проводником. Мигрирующие ионы, так же как и ионы хлора, называются в данном случае сопровождающими ионами. Знак их электрического заряда противоположен знаку заряда противоионов. [35]
Проведенные исследования позволили предположить, что первичный механизм биологического действия препарата связан с увеличением проводимости и нарушением структурной организации клеточных мембран. При этом увеличение проводимости мембран обусловлено липофильностью мембранно-активной формы картоцида. [36]
При оценке изменений сопротивление мембраны нейрона следует рассмотреть два случая. Первый сводится к неселективному изменению проводимости мембраны. Второй случай более сложный. Под влиянием инъецируемого тока возрастает проводимость к тому типу ионов, пассивное движение которых по электрохимическому градиенту создает потенциал, противоположный по знаку тому потенциалу, который был создан инъекцией тока. Рассмотрим в качестве примера инъекцию катионов, создающую деполяризационный сдвиг. Под влиянием инъекции катионов в ряде нейронов моллюсков происходит повышение калиевой проводимости. Выход калия вызывает гиперполяризацию нейрона, компенсирующую деполяризационный сдвиг. При выключении инъецируемого тока повышенная калиевая проводимость и продолжающийся выход калия из клетки создают смещение МП в сторону гиперполяризации, которая может длиться достаточно долго. [37]
Другая модель - модель эстафетного переносчика - предполагает, что переносчики неподвижны и образуют цепи, расположенные поперек мембраны, а катионы пересекают мембрану вдоль этих цепей, перескакивая от одного переносчика Т к другому. Обе модели приводят к колоколооб-разной зависимости проводимости мембран на постоянном токе от концентрации ионов. Однако при малом содержании С в растворе и больших концентрациях Т в мембране проводимость ее по переменному току высокой частоты мала для эстафетного механизма и значительна при механизме подвижных переносчиков. [38]
Другая модель - модель эстафетного переносчика - предполагает, что переносчики неподвижны и образуют цепи, расположенные поперек мембраны, а катионы пересекают мембрану вдоль этих цепей, перескакивая от одного переносчика Т - к другому. Обе модели приводят к колоколообразной зависимости проводимости мембран по постоянному току от концентрации ионов. Однако при малом содержании С в растворе и больших концентрациях Т - в мембране проводимость ее по переменному току высокой частоты мала для эстафетного механизма и значительна при механизме подвижных переносчиков. [39]
Поскольку концентрации ионов в мембране зависят от их концентраций во внешнем растворе ( или растворах), с которым мембрана находится в соприкосновении, проводимость мембран возрастает с увеличением концентрации внешнего раствора и наоборот. Таким образом, можно ожидать, что проводимость мембраны имеет низший предел, который служит характеристикой данной мембраны при малых концентрациях внешнего раствора. [40]

Глицин проявляет примерно такое же ингибиторное действие, как GABA. Он стабилизирует потенциал покоя, повышая С1 - проводимость синаптической мембраны. Обе эти аминокислоты анатомически локализованы в различных, но иногда перекрывающихся отделах центральной нервной системы. Глицин - это, по-видимому, основной ингибиторный медиатор спинного мозга и ствола головного мозга, тогда как GABA чаще находят в мозге. Хлорные каналы этих двух рецепторов тоже, вероятно, не идентичны. Конвульсант стрихнин, неконкурентный антагонист глицинового синапса, взаимодействует непосредственно с его хлорными каналами ( гл. [42]
В связи с изложенным представляет интерес работа [56], в которой экспериментально показано, что при установлении равновесия ( V87) газообразные ислород и сера могут перемещаться через силикатную мембрану против градиента их концентраций. Действительно, если равновесие определяется только реакцией (V.87) и проводимость мембраны чисто ионная, то равновесие по обе ее стороны зависит не от отдельных парциальных давлений S2 и О2, а от их отношения. [43]
Подвижные ионы любого заряда имеют в фазе мембраны подвижность, значительно меньшую, чем во внешнем растворе; это, возможно, вызвано совместным действием электростатических и междуионных сил притяжения, существующих в мембране. Отсюда следует ожидать, что при умеренных и высоких концентрациях внешнего раствора проводимость мембраны будет меньше его проводимости. Как уже отмечалось, при малых внешних концентрациях проводимость мембраны приближается к минимуму, проводимость же внешнего раствора при бесконечном разведении достигает нуля. При некотором значении концентрации внешнего раствора эти две проводимости становятся равными и создают впечатление гомогенности системы. [44]
Со времени открытия более полувека тому назад аце-тилхолина арсенал трансмиттеров пополнился примерно десятком других органических соединений, выступающих в качестве синаптических веществ-передатчиков. Они накапливаются в синаптических пузырьках, скользят к пост-синаптической мембране на входе нервной клетки, где соединяются с протеиновыми рецепторами и на короткое время селективно изменяют проводимость мембраны для определенных ионов. Длящиеся от 10 до 20 миллисекунд постсинаптические де - и гиперполяризации в несколько милливольт мембранной поляризации ( внутриклеточная - около 80 милливольт по отношению к внеклеточной) суммируются по времени и пространству и ведут к дальнейшему возбуждению в форме акциональных потенциалов, если не будет повышен критический порог поляризации ( например, в 65 милливольт) у аксонного выступа. Если темпоральная суммация локальных возбуждающих постсинаптических потенциалов ( ВПСП) и тормозящих постсинаптических потенциалов ( ТПСП) является следствием сукцессивных ( наследственных) синаптических возбуждений, то локальная суммация зависит от одновременного поступления потенциалов через различные контактирующие участки. [45]
Страницы: 1 2 3 4