Проназа
Cтраница 2
Исследование, проведенное с использованием в качестве субстратов различных синтетических нуклеотидо - ( Р - - Ы) - аминокислот ( пептидов), показало, что гидролитические ферменты, расщепляющие пептидные или фосфодиэфирные связи ( проназа, панкреатическая РНК-аза, фосфодиэстераза), не расщепляют фосфоамидную связь в этих соединениях. [16]
При более высокой температуре ( 30 - 35) действие пепсина, фи-цина и проназы приводит к большему или меньшему разрушению тройной спирали макромолекулы коллагена, связанному, по-видимому, с гидролизом пептидных связей. Проназа даже при 20 вызывает укорачивание молекулы коллагена с ее А-конца приблизительно на 5 %, после чего молекула становится особенно чувствительной к действию других протеаз. Действие трипсина на коллаген при всех условиях крайне ограничено. [17]
Как видно из приведенных в обзоре данных, для биосинтеза внеклеточных протеиназ с целью получения препаратов ферментов наиболее часто используют штаммы, относящиеся к разным видам рода Actinomyces. Такие препараты, как проназа, протелин, протеиназы Str. Ионы кальция стабилизируют все препараты, кроме протофрадина. [18]
Сиало вые кислоты, входящие в состав гликопротеинов, защищают их от действия протеолитических ферментов, поэтому необходимо предварительно отщепить их нейраминидазой. После удаления сиаловых кислот гликопротеины частично расщепляются протеолитическими ферментами ( папаин, проназа) на глнкопептиды, доступные для дальнейшего исследования. Гликопептиды затем обрабатывают ферментами, отщепляющими аминокислоты ( например, карбоксипештидазой после защиты конечной аминогруппы), и превращают в олигосахариды, содержащие только одну аминокислоту. Углеводные компоненты гликопротеинов, имеющих щелочелабильную связь углевод-аминокислота, отщепляют щелочной обработкой или ферментами, разрушающими связи такого типа. Полученные таким образом тетеросахариды или гете-росахариды с одной аминокислотой очищают от примесей пептидов и аминокислот переосаждением из водноорганических смесей ( вода - спирт, вода-ацетон), экстракцией фенолом, хроматографией, электрофорезом или гель-фильтрацией. [19]
К этой группе ферментных препаратов относят: во-первых, протеиназы как животного, так и микробного, грибкового и растительного происхождения ( трипсин, хипотрипсин, эластаза, проназа, коллагеназа, папаин, бро-мелин, фицин, дикиназа и др.), обладающие высокой способностью гидролизовать денатурированные массы белков, разжижать вязкие и трудноотделяемые экссудаты, снижать антигенные свойства ряда токсинов; во-вторых, деполимеразы нуклеиновых кислот и нуклеопротеидов - ДНК-азы и РНК-азы, использующиеся при лечении больных хроническими гнойными заболеваниями легких и при других нагноительных процессах ( конъюнктивиты, кератиты и пр. [20]
Инактивация связана с фосфорилированием серина в активном центре фермента. Исходя из количества фосфора, связанного трипсином проназы, и молекулярного веса проназы, приблизительно равного 20 000, Вальби [164] рассчитал, что на долю трипсиноподобной протеиназы приходится около 26 % общего количества белка проназы. [21]
Необходимо отметить, что между протелином и проназой существует большое сходство. Это сходство проявляется в высокой аротеоли-тической активности препаратов, в близких рН - оптимумах активности и стабильности этих препаратов, в числе и специфичности содержащихся в них протеиназ. Протелин подобен проназе также по глубине гидролиза различных белков и по стабильности в растворах мочевины. [22]
Ферментативные методы гидролиза основаны на избирательности действия протеолитических ( вызывающих распад белков) ферментов, расщепляющих пептидные связи, образованные определенными аминокислотами. В частности, пепсин ускоряет гидролиз связей, образованных остатками фенилаланина, тирозина и глутаминовой кислоты, трипсин - аргинина и лизина, химотрипсин-триптофана, тирозина и фенилаланина. Ряд других ферментов, например папаин, субтилизин, проназа и другие бактериальные протеиназы, также используется для неполного гидролиза белков. В результате полипептидная цепь расщепляется на мелкие пептиды, содержащие иногда всего несколько аминокислот, которые отделяют друг от друга сочетанными электрофоретическими и хроматографическими методами, получая своеобразные пептидные карты. Далее определяют чередование аминокислот в каждом индивидуальном пептиде. Завершается работа воссозданием первичной структуры полной полипептидной цепи на основании определения последовательности аминокислот в отдельных пептидах. [23]
Среди них наиболее предпочтительными благодаря своей селективности являются ферментативные. С помощью протеаз, например, если действовать ими сначала на наружную, а затем на внутреннюю поверхности мембраны, можно определить, различна ли структура и функция белка на разных сторонах бислоя. Было показано, в частности, что при перфузии аксона проназой, мишенью действия фермента оказались белки, участвующие в инактивации натриевого канала, и, следовательно, они должны быть размещены на внутренней стороне мембраны. [24]
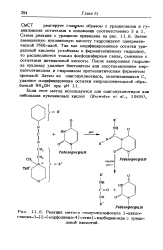 |
Реакция мето-п-толуолсульфоната 1-цикле - гексил-3 - [ 2 - ( морфолинил-4 - этил ] - карбодиимида с уриди. [25] |
Схема реакции с урацилом приведена на рис. 11.8. Затем замещенную нуклеиновую кислоту гидролизуют панкреатической РНК-азой. Так как модифицированные остатки ури - диловой кислоты устойчивы к ферментативному гидролизу, то расщепляются только фосфодиэфирные связи, смежные с остатками цитидиловой кислоты. После завершения гидролиза нуклеазу удаляют бентонитом или восстанавлением мер-каптоэтанолом и гидролизом протеолитическим ферментом проназой. [26]
Протеолитическая активность довольно четко выражена у большого числа видов и штаммов актиномицетов. Однако протеолитические ферменты актиномицетов привлекли внимание исследователей только в 50 - е годы, после того, как эти микроорганизмы стали культивировать с целью получения антибиотиков. Несколько позднее были разработаны методы раздельного выделения продуктов биосинтеза и получены такие препараты, как проназа и протелин из Streptomyces griseus и римопротелин из Act. [27]
Наиболее высокая кол-лагеназная активность обнаружена во фракциях 8 и 9 содержащих щелочные протеазы проназы. Это свидетельствует об ограниченности действия протеаз проназы на Коллаген. [28]
Известно, что каждый фермент имеет свой характерный температурный оптимум, определяемый двумя противоположно идущими процессами - обычным повышением скорости реакции от нагревания и денатурацией, ускоряющейся по той же причине. Температурный оптимум может быть повышен, если ферменты стабилизировать; тогда все катализируемые процессы возможно будет значительно ускорять. Даже небольшое увеличение устойчивости фермента к нагреванию может быть весьма важным, особенно если при этом удастся превысить температуры, обычно переносимые микробами. Таковы, например, папаин, проназа, рибонуклеаза и др.; б) выделение промышленных термоустойчивых ферментов из термофильных микроорганизмов. [29]
Страницы: 1 2