Кооперативный процесс
Cтраница 1
 |
Схема многоступенчатого механизма возбуждения антястоксовской люминесценции. [1] |
Кооперативные процессы заключаются в таком взаимодействии нескольких возбужденных ионов, при котором энергия, запасенная ими, в едином акте передается одному иону, переводя его в возбужденное состояние. [2]
Понятие кооперативных процессов, введенное применительно к фазовым превращениям в ансамблях, содержащих большое число частиц, трактуется в современной биофизике расширительно. Любые явления, определяемые взаимодействием даже малого числа элементов системы, принято называть кооперативными. [3]
Возникновение кооперативных процессов, как и формирование и развитие новых структур, непосредственно связано с действием случайных факторов. Мысль о том, что без случайного невозможно появление нового, высказанная в форме догадки еще античными философами Демокритом и Лукрецием Каром, нашла блестящее подтверждение в синергетике. Действительно, началом любого развития являются случайные изменения, которые постепенно приводят к неустойчивости системы. В результате взаимодействия большого числа случайных факторов в открытых неравновесных системах происходит их взаимное согласование и возникают кооперативные процессы, сопровождающиеся коллективным поведением элементов вновь образующейся структуры. По какому пути пойдет дальнейшая эволюция, какая альтернатива будет выбрана системой, во многом также зависит от случайных факторов. Именно с ними в существенной мере связано появление нового в развитии систем, в частности социально-экономических. [4]
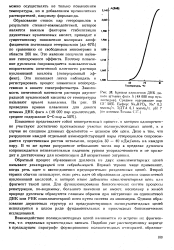 |
Кривая плавления ДНК дикого штамма фага А ( 48 000 пар нук-леотидов. Среднее содержание пар GC 50 %. Буфер. Na2HP04, Na 0 2 ммоль / л, ЭДТА 0 1 ммоль / л. рН 7 ( по данным Лохова С.Г. [5] |
Плавление представляет собой кооперативный процесс, в котором одновременно участвуют достаточно протяженные участки полинуклеотидиых цепей, а в случае не слишком длинных фрагментов - целиком обе цепи. Дело в том, что разрушение каждой отдельной взаимодействующей пары гетероциклов сопровождается существенным ростом энтальпии, порядка 30 - 40 кДж / моль на каждую пару. В то же время разрушение небольшого числа пар в пределах дуплекса сопровождается незначительным падением уровня упорядоченности и не приводит к достаточному для компенсации АН возрастанию энтропии. [6]
Перенос протона посредством кооперативного процесса, описанного уравнением (12.49), известен под названием проводимости Грот-гуса, наблюдаемой в протолитических реакциях в водном растворе. [7]
Значит, при кооперативном процессе высота актива ционного барьера не есть величина постоянная, она зависит от числа объектов, уже переваливших через этот барьер. [8]
Причиной его является не кооперативный процесс, а стремление к равновесию системы внедренных атомов, находящихся во внешнем по отношению к ней поле решетки узлов. [9]
В отличие от термодинамики кооперативных процессов, их кинетика разработана совершенно недостаточно. В сущности физика пока не располагает полной кинетической теорией кристаллизации или плавления. [10]
Осаждение макромолекул из раствора представляет собой кооперативный процесс. Необходимо найти возможность превратить с помощью какого-либо функционального преобразования кривую распределения по молекулярным весам С ( М) в кривую изменения мутности с ( у) и наоборот. Другими словами, количество вещества dc ( у), выпавшее в осадок в диапазоне между у и у dy, будет зависеть ( в различной степени) от концентрации макромолекул всех размеров в данном образце и, в частности, от концентрации всех макромолекул, еще находящихся в растворе. Теории, которая была бы основана на этом общем положении и могла бы явиться исходной для расчета кривых изменения мутности, пока не существует. [11]

С) и отличается чертами кооперативного процесса, напоминающего фазовый переход. По достижении томп-ры плавления резко падают вязкость раствора и его онтич. Наличие определенного диапазона темп-р, в к-ром развивается денатурация, приписывают гетерогенности ДНК. [13]

Сегодня мы знаем, что кооперативными процессами являются как связывание агониста с рецептором, так и последующая стадия открывания ионного канала. [15]
Страницы: 1 2 3 4