Путь - окисление
Cтраница 3
Вышеуказанные пути образования а превращений пентозофосфа-тов аналогичны тем, которые имеют место в адотомическом пути окисления глшозы при дыхании ( пентозофосфатный цикл), но направлены в противоположную сторону, поэтому цикл Кальвина называют восстановительным пентозофосфатным циклом. [31]
Из утого выражения видно, что отношение выходов формальдегида и ацильных перекисей и вероятность формальдегидного пути окисления будут возрастать с ростом температуры, с уменьшением давления и концентрации кислорода. [32]
Пути окисления главных жирных кислот с разветвленной цепью - изобу-тирата, а-метилбутирата и изовалерата - очень сходны с путями окисления валина, изолейцина и лейцина соответственно. Эти процессы рассматриваются в гл. [33]
В случае 4-метил - 2 6-грет-бутилфенола, который часто используют в качестве добавки к нефти и смазочным маслам, установлено два пути окисления. [34]
При высоких температурах сварочного процесса окислительная способность углекислого газа увеличивается, главным образом вследствие образования атомарного кислорода, и начинает преобладать второй путь окисления металла. [35]
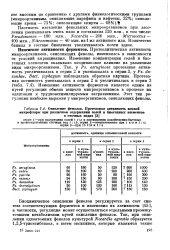 |
Окисление фенолов. Протеазная активность иловой. [36] |
Биохимическое окисление фенолов регулируется за счет синтеза соответствующих ферментов и изменения их активности 115 ], в частности, регуляция может осуществляться отдельными промежуточными метаболитами путей окисления фенолов. [37]
Одни исследователи считают, например, что в организме человека и животных около 60 % глюкозы окисляется апотомическим путем, другие, напротив, приходят к выводу, что этот путь окисления не имеет в целостном организме практического значения. Такое расхождение во взглядах объясняется большими трудностями, возникавшими до последнего времени при постановке и особенно при истолковании получаемых экспериментальных данных. [38]
Следует иметь в виду, что при добавлении а-кетоглутаровой кислоты как субстрата окисления необходимо вносить в пробу также и малоновую кислоту, которая, как известно, является ингибитором сукциндегидразы и, следовательно, затормаживает быстрый путь окисления янтарной кислоты, образующейся при окислении а-кетоглутаровой кислоты, и не позволяет исказить результатов окисления а-кетоглутаровой кислоты. [39]
Хотя для внесения полной ясности в этот вопрос необходимы еще дополнительные исследования, однако уже и сейчас можно думать, что в ряде тканей ( надпочечники, селезенка, зобная железа, лимфатические узлы, лактирующая молочная железа и др.) апотомический ( гексозомонофосфатный) путь окисления глюкозы занимает достаточно видное место. [40]
Этот путь окисления парафинов всегда связан с деструкцией углерод-углеродных связей. Процесс протекает в жидкой фазе - термически или в присутствии катализатора при температуре от 105 - 120 до 170 - 200 С. [41]
Этот путь окисления парафинов всегда связан с деструкцией углерод-углеродных язей. Процесс протекает в жидкой фазе - термически или в присутствии катализатора при температуре от 105 - 120 до 170 - 200 С. [42]
Открытие пути прямого окисления углеводов, или, как его называют, пентозофосфатного цикла, принадлежит О. Расхождение путей окисления углеводов-классического ( цикл трикарбоновых кислот, или цикл Кребса) и пентозофосфатного-начинается со стадии образования гексозомонофосфата. Если глюкозо-6 - фосфат изомеризуется во фруктозо-6 - фосфат, который фосфо-рилируется второй раз и превращается во фруктозе-1 6-бисфосфат, то в этом случае дальнейший распад углеводов происходит по обычному гликолитическому пути с образованием пировиноградной кислоты, которая, окисляясь до ацетил - КоА, затем сгорает в цикле Кребса. [43]
Представление о существовании в тканях животных, растений и микробов иного механизма окисления углеводов, отличающегося от рассмотренного выше, были развиты на основе работ главным образом Варбурга, Диккенса и В. А. Энгельгардта, показавших возможность окисления гексо-зомонофосфата в клетках животных ( например, в эритроцитах) без предварительного расщепления на две фосфотриозы. Расхождение путей окисления углеводов - классического, при участии лимоннокислого цикла Кребса, и пентозного - начинается на стадии образования гексозомонофосфата. [44]
Следует подчеркнуть, что существовавшее ранее представление о пути нефосфорилирующего окисления как характерном главным образом для патологического обмена в настоящее время оставлено. Однако в митохондриях кровяных пластинок обнаружен лишь путь окисления, не сопряженного с фосфорилированием, и добавление к реакционной среде таких агентов, как система гексокиназа - глюкоза, АТФ, ЭДТА, ДПН, не приводило к выявлению сопряжения окисления с фосфорилированием. [45]
Страницы: 1 2 3 4