Пентозофосфатный путь
Cтраница 4
Центральную роль в превращениях глюкозы и саморегуляции углеводного обмена в печени играет глюкозо-6 - фосфат. Он резко тормозит фосфоролитическое расщепление гликогена, активирует ферментативный перенос глюкозы с уридиндифосфоглюкозы на молекулу синтезирующегося гликогена, является субстратом для дальнейших гликолитических превращений, а также окисления глюкозы, в том числе по пентозофосфатному пути. [46]
В организме человека образуется 160 млн эритроцитов в минуту; они циркулируют в крови 110 - 120 дней и затем разрушаются. Зрелые эритроциты - это безъядерные клетки, не содержащие мРНК, рибосом и митохондрий; основным энергетическим процессом в них является гликолиз. В зрелом эритроците активен пентозофосфатный путь, в процессе которого происходит восстановление НАДФН. Синтез гемоглобина происходит в процессе развития эритроцитов из стволовых клеток костного мозга, на стадии образования незрелой безъядерной красной клетки - ретикулоцита, которая поступает в кровь. Ретикулоциты содержат много глобиновой мРНК и активно синтезируют гемоглобин, пока клетка в процессе созревания не превращается в эритроцит, утрачивающий мРНК, рибосомы и митохондрии. В результате зрелый эритроцит обладает упрощенным метаболизмом, нацеленным на сохранение целостности мембраны и предотвращение окисления гемоглобина. Эритропо-эз стимулируется белком эритропоэтином, вырабатываемым в мозговом слое почек. [47]
Некоторые цианобактерий образуют специализированные клетки с утолщенной клеточной стенкой - гетероцисты - основное место азотфиксации. Здесь отсутствует ФС II, а в качестве восстановителей используются сахара, транспортируемые через плаз-модесмы из соседних клеток. В гетероцистах не функционирует цикл Кальвина, но имеется пентозофосфатный путь. У Gloeothece азотфиксация идет в обычных вегетативных клетках, в которых также нет ФС II. Некоторые безгетеро-цистные цианобактерии фиксируют азот в обычных клетках, но только в анаэробных условиях. [48]
Катаболические процессы являются в основном окислительными и служат донорами высокоэнергетических электронов, для анаболизма же характерно обратное. Основным донором электронов в восстановительных реакциях биосинтеза является НАДФН, восстановление которого происходит в реакциях катаболизма, большей частью в пентозофосфатном пути окисления глюкозы. [49]
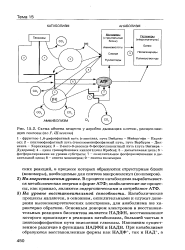 |
Схема обмена веществ у аэробно дышащих клеток, расщепляющих гексозы ( по Г. Шлегелю. [50] |
Катаболические процессы являются, в основном, окислительными и служат донорами высокоэвергетических электронов, для анаболизма же характерно обратное. Основным донором электронов в восстановительных реакциях биосинтеза является НАДФН, восстановление которого происходит в реакциях катаболизма, большей частью в пентозофосфатном пути окисления глюкозы. [51]
Углеводы пентозофосфатного цикла - триозофосфаты, например соединение ( 6), и 3-фосфоглицериновая кислота ( 5) - являются не только важными промежуточными соединениями в фотосинтетическом связывании диоксида углерода, но и ключевыми соединениями в промежуточном метаболизме углеводов во всех организмах. Метаболизм глюкозы и других углеводов осуществляется по хорошо известному гликолитическому пути; они участвуют также в цикле трикарбоновых кислот; таким путем происходит превращение большей части углеводов в большинстве организмов. Альтернативным путем окисления углеводов, в количественном отношении, правда, менее важном, чем гликолиз и цикл трикарбоновых кислот, является пентозофосфатный путь. Его важность для организмов определяется тем обстоятельством, что он служит источником NADPH и пентозофосфатов. D-глюконолактон-б - фосфат до О-рибулозо - 5-фосфата; при этом образуется также ADPH. Пенто-зофосфат затем превращается в фосфаты других Сахаров, например D-фруктозо-б - фосфат и D-глицеральдегид - З - фосфат, которые затем включаются в метаболизм по гликолитическому пути, а также в Л - эритрозо-4 - фосфат, который наряду с фосфоенолпиру-ватом является исходным соединением в биосинтезе ароматических аминокислот через шикимовую кислоту. [52]
Страницы: 1 2 3 4