Клонирующий вектор
Cтраница 2
В этой системе на неонкогенной Ti-плазмиде синтезируются продукты vir - генов, которые мобилизуют участок Т - ДНК бинарного клонирующего вектора. Продуцируя белки, кодируемые v / r - генами, неонкогенная Ti-плазмида выступает в роли помощника, способствуя встраиванию Т - ДНК из бинарного клонирующего вектора в хромосомную ДНК растения. [16]
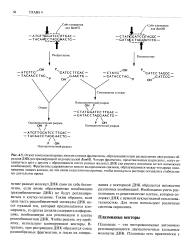
ДНК само по себе бесполезно, если вновь образованные комбинации ( рекомбинантные ДНК) не будут реплицироваться в клетке-хозяине. Таким образом, если одна часть рекомбинантной молекулы ДНК несет нужный ген, который предполагается клонировать, то другая должна содержать информацию, необходимую для репликации в клетке рекомбинантной ДНК. Чтобы решить эту проблему, используют клонирующие векторы. [18]
Синтетический двухцепочечный олигонуклеотид с одним тупым концом и одним липким. После пришивания адаптера тупым концом к ДНК-мишени последнюю можно встраивать в подходящий вектор, используя приобретенный ею липкий конец. Синтетический одноцепочечный олигонуклеотид, у которого после самогибридизации появляются липкие концы и внутренний сайт для рестрици-рующей эндонуклеазы. Когда адаптер встраивают в клонирующий вектор, у последнего появляется новый сайт рестрикции. [19]
Во втором случае используют коинтегративную векторную систему. Коинтегративный вектор и неонкогенная Ti-плазмида-помощник содержат гомологичные последовательности, которые образуют сайт для гомологичной рекомбинации in vivo. Обычно эти последовательности расположены в Т - ДНК. После рекомбинации клонирующий вектор становится частью неонкогенной Ti-плазми-ды, которая содержит v / r - гены, необходимые для переноса Т - ДНК в растительную хозяйскую клетку. [20]
Страницы: 1 2