Раздражение - нерв
Cтраница 1
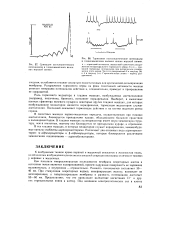 |
Суммация постсинаптических потенциалов в гладкомышечных волокнах морской свинки.| Тормозные постсинаптические потенциалы в гладкомышечном волокне кишки морской свинки. [1] |
Раздражение тормозного нерва на фоне спонтанной активности мышцы угнетает генерацию потенциалов действия и, следовательно, приводит к прекращению ее сокращений. [2]
 |
Возможные положения ортодромного импульса в момент t О. [3] |
При раздражении нерва двумя импульсами с переменной задержкой времени Т второго импульса относительно первого. [4]
При раздражении нерва теплообразование значительно возрастает. Так же как и в мышце, тепло выделяется в две фазы, которые обозначаются как начальное и запаздывающее теплообразование. [5]
 |
Эквивалентная электрическая схема тормозного синаптического входа.| Тормозной постсинаптический потенциал ( ТПСП в мотонейронах спинного мозга кошки. [6] |
ТПСП, вызванный раздражением нерва; б - ТПСП, вызванный раздражением вставочного нейрона. [7]
 |
Влияние ингибитора холинэстеразы ( нео-стигмина на длительность постсинаптического потенциала ( потенциала концевой пластинки одиночного мышечного волокна.| Оптимум ( Ор и пессимум ( Ps. [8] |
При действии ингибитора холинэстеразы ритмическое раздражение нерва вызывает выраженную суммацию ПКП, что ведет к стойкой деполяризации постсинаптической мембраны и блоку проведения импульсов с нервного волокна на мышечное. [9]
Следует отметить, что и в отсутствие ингибиторов холинэстеразы при условии очень частого раздражения нерва постсинаптические потенциалы ( ПКП), вызываемые каждым нервным импульсом, суммируются, поскольку в межимпульсный интервал холинэстераза не успевает полностью расщепить выделяющийся в нервном окончании ацетилхолин. В результате суммации потенциалов постсинаптическая мембрана все более и более деполяризуется. [10]
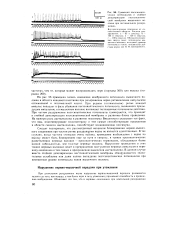 |
Суммация постсинапти-ческих потенциалов и стойкая деполяризация постсинаптиче-ской мембраны мышечного волокна при пессимальном раздражении. [11] |
Следует подчеркнуть, что рассмотренный механизм блокирования нервно-мышечного соединения при частом ритме раздражения нерва не является единственным. В тех случаях, когда частота стимулов очень высока, проведение возбуждения с нерва на мышцу может быть блокировано еще на пути к синапсу, в тонких пресинаптических разветвлениях нервных волокон - пресинаптических терминалях, обладающих более низкой лабильностью, чем толстые нервные волокна. Нарушение проведения в этих тонких нервных волокнах ведет к прекращению поступления нервных импульсов к нервному окончанию и тем самым к прекращению выделения ацетилхолина. В данном случае вместо стойкой деполяризации постсинаптической мембраны обнаруживается значительное ослабление или даже полное выпадение постсинаптических потенциалов при неизменном уровне потенциала покоя мышечного волокна. [12]
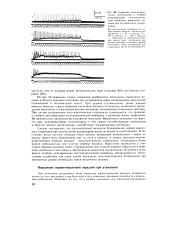 |
Суммация постсинапти-ческих потенциалов и стойкая деполяризация постсинаптиче-ской мембраны мышечного волокна при пессимальном раздражении. [13] |
На рис. 56 приведена запись изменения мембранного потенциала мышечного волокна в области концевой пластинки при раздражении нерва ритмическими импульсами оптимальной и пессимальной частот. При редком ( оптимальном) ритме каждый импульс попадает в фазу убывания постсинаптического потенциала, вызванного предыдущим импульсом, и в мышечном волокне возникают полноценные потенциалы действия. При частом раздражении постсинаптические потенциалы суммируются, что приводит к стойкой деполяризации постсинаптической мембраны и развитию блока проведения. На важную роль ацетилхолина в развитии пессимума Введенского указывает тот факт, что яды, инактивирующие холинэстеразу и тем самым способствующие накоплению в области синапса ацетилхолина, способствуют возникновению пессимума. [14]
В некоторых идентифицированных нейронах ответ, вызванный апликацией АХ, точно воспроизводит эффект кожного раздражения или ортодромного раздражения нерва. В ряде случаев медиатор действует на сому нейрона непосредственно как нейрогуморальный фактор. В некоторых нейронах на естественные раздражения регистрируются гигантские ВПСП или ТПСП, воспроизводящие форму ответа на прямую апликацию медиатора. В синаптических нейронах такие потенциалы могут включать ПД синаптического происхождения. В латентных пейсмекерных нейронах гигантский ВПСП при достижении условий запуска пейсмекерных потенциалов приводит к генерации пейсмекерных ПД. В актуальном пейсмекерном нейроне при гигантском ВПСП происходит изменение частоты пейсмекерных потенциалов и связанных с ним ПД. [15]
Страницы: 1 2 3 4