Распределение - аминокислота
Cтраница 1
Распределение аминокислот и других важных аминосоединений показано на фиг. В 18 случаях нанесенные на бумагу аминокислоты обнаруживались при спектрофотометрическом определении при помощи модифицированного нингидринового реактива ( см. стр. Для глицина и серина после хро-матографирования в обратном направлении вырезанного участка хроматограммы, проявленной растворителем П - Э - ФФ, были получены фотометрические отсчеты, составляющие соответственно 82 и 75 % величин, найденных при прямом анализе образцов. [1]
Распределение аминокислот между форменными элементами и окружающей их плазмой неравномерно. В эритроцитах содержится всегда больше аминокислот, чем в окружающей их плазме. [2]
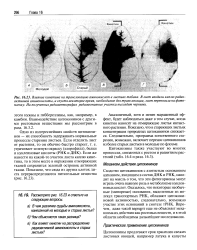
Как влияет кинетин на распределение радиоактивной аминокислоты в старых листьях. [4]

Как следует из таблицы, распределение аминокислот по четырем вертикальным столбцам отражает тенденцию к дискриминации одних и предпочтительному включению других. Аминокислоты, расположенные в крайнем левом столбце, могут только дискриминироваться, а в крайнем правом - только накапливаться. Аминокислоты, занимающие промежуточное положение, могут и дискриминироваться, и накапливаться, причем для аминокислот правой половины таблицы больше вероятность накопления. [6]
Эти свойства коллидина и вызывают распределение аминокислот через весь лист бумаги на двухмерной хроматограмме. Нами была подобрана смесь пиридина ( или а-пиколина), хинолина и воды, удовлетворяющая основным свойствам коллидина. Такая смесь может заменять коллидин в качестве второго растворителя для двухмерной хроматографии, а также использоваться для некоторых целей для одномерной хроматографии. Состав смесей: 1) на 100 см3 смеси из 70 см3 пиколина, 11 см3 хинолина и 19 см3 воды; 2) 66 см3 пиридина, 17 см3 хинолина, 17 см3 воды. [7]
Изложение материалов, связанных с распределением аминокислоты, было начато с качественных представлений о протекающих в этой системе процессах и устанавливающихся в ней равновесиях. [8]
Как видно из табл. 5.2, распределение аминокислот по кодонам весьма неравномерно. Трем аминокислотам - лейцину, серину и аргинину - соответствует по шесть кодонов, пяти аминокислотам - глицину, аланину, валину, пролину и треонину - по четыре, изолейцину - три кодона, лизину, аспартату, аспараги-ну, глутамату, глутамину, фенилаланину, тирозину, гйстидину и цистеину - по два, а метионину и триптофану - по одному кодону. Три кодона - UAA, UAG и UGA - не соответствуют ни одной из аминокислот. Они являются сигналами для прекращения синтеза полипептидной цепи: их называют кодонами-терминатора-ми. В восьми случаях природа кодируемой аминокислоты однозначно определяется первыми двумя нуклеотидами кодона: UC - серии, АС - треонин, CU - лейцин, GU - валин, СС - пролин, CG - аргинин, GC - аланин и GG - глицин. [9]
Экспериментально определяют степень экстракции или коэффициент распределения аминокислоты при разных рН воднои фазы, затем по уравнению ( 54) или ( 55) вычисляют константу распределения. [10]
Экспериментально определяют степень экстракции или коэффициент распределения аминокислоты при разных рН водной фазы, затем по уравнению ( 51) или ( 52) вычисляют константу распределения. [11]
На основании результатов хроматографирования на бумаге вычерчивают диаграмму, показывающую распределение аминокислот в собранных фракциях [ см. Partridge, Brimley, Biochem. Высыпают смолу в банку, а колонку моют и высушивают. [12]
В настоящее время некоторыми авторами высказывается идея о том, что распределение полярных и неполярных аминокислот вдоль полипептидной цепи является одним из важных элементов кодирования пространственной структуры глобулярных белков. Еще Фишером [55] было показано, что соотношение суммарных объемов полярных и неполярных аминокислотных остатков может обусловливать форму белковой молекулы ( сферическую или вытянутую), а также способность образовывать четвертичные структуры. [13]

Разделение сложной смеси аминокислот на составные части основано на различии коэффициентов распределения аминокислот в двух несмешивающихся растворителях. [15]
Страницы: 1 2