Метаболическая реакция
Cтраница 1
Окислительные метаболические реакции в ряду кадалина часто приводят к ароматизации одного или обоих колец. Соединения с одним ароматическим кольцом, простой пример которых - углеводород калакорен 2.227, выделяют в подгруппу каламена, Каламеновый непредельный кетон лацини-лен С 2.228 выполняет функцию фитоалексина у хлопкового растения. В большом количестве в хлопке содержится также димерное нафталиновое производное кадалина госсипол 2.229. Он известен науке с конца XIX столетия, но только недавно выявлена его ценность как биологически активного вещества. Перечень физиологических эффектов госсипола весьма велик. Что же касается практического использования, то он нашел применение при лечении хронического гломерулонефрита и аллергических кожных болезней. [1]
Метаболические реакции адреналина изучены in vitro на тканевых препаратах. Метилирование адреналина происходит в некоторых тканях; у крысы активность этого процесса в различных тканях убывает в следующем порядке: печень почки селезенка тонкий кишечник легкие мозг - сердечная мышца. В скелетной мышце метилирования адреналина не обнаружено. Метилирование происходит в печени ряда видов млекопитающих, включая и человека [40]; фермент локализован в растворимой фракции гомогенатов печени. Метилирование, очевидно, является основным путем инактивации адреналина и норадреналина, так как 3 - О-метиловые эфиры полностью лишены физиологической активности, присущей исходным соединениям. Моноаминооксидаза осуществляет превращение метилированных гормонов в 4-окси - З - метоксифенилгликольальдегид, который затем окисляется и восстанавливается ( см. стр. [2]
Большинство конкретных метаболических реакций также демонстрирует отчетливую периодичность на протяжении суток. Уже упоминалось о закономерных суточных движениях листьев и цветков. Столь же закономерно сменяют друг друга в суточном цикле фотосинтез и дыхание. Суточная периодика свойственна и ряду других физиологических процессов у растений. [3]
Обычно это метаболические реакции, связанные с окислением определенного субстрата ( биохимического топлива) или с разрывом богатых энергией ( макроэрги ческих) полифосфатных связей в молекуле аде-нозинтрифосфата ( АТР) с образованием аденозиндифос-фата ( ADP) и иона фосфата. [4]
Почти все метаболические реакции связаны между собой, так как продукт одной ферментативной реакции служит субстратом другой, которая является следующим этапом данного процесса. [5]
Практически все метаболические реакции катализируются соответствующими ферментными системами, большая часть из которых локализована в эндо-плазматическом ретикулуме. Следует помнить, что ферменты, изменяющие структуру ксенобиотиков, сами подвержены действию чужеродных соединений. [6]

Почти все метаболические реакции в конечном счете связаны между собой, поскольку продукт одной ферментативной реакции служит субстратом для другой, которая в данном процессе играет роль следующей стадии. [8]
Стереоспецифичность многих метаболических реакций может объясняться причинами, имеющими отношение к формальным механизмам катализа. Например, одинаковая стереохимическая специфичность четырех кето-альдегид-изомераз привела Розе [13] к предположению об общем г с-эндиольном механизме для этих четырех ферментов и об одновременном разрыве связей в каждой из этих реакций. [9]
Систематическое изучение метаболических реакций составляет одну из главных задач биохимии. [10]
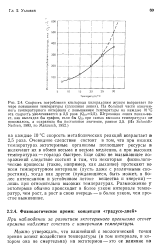
С скорость метаболических реакций возрастает в 2 5 раза. Очевидное следствие состоит в том, что при низких температурах эктотермные организмы поглощают ресурсы и включают их в обмен весьма и весьма медленно, а при высоких температурах - гораздо быстрее. Еще одно не вызывающее возражений следствие состоит в том, что некоторые физиологические процессы ( такие, например, как дыхание) протекают во всем температурном интервале ( пусть даже с различными скоростями), тогда как другие ( нуждающиеся, быть может, в более интенсивном и устойчивом потоке вещества и энергии) - лишь при относительно высоких температурах. Размножение у эктотермов обычно происходит в более узком интервале температур, чем рост, а рост в свою очередь - в более узком, чем простое выживание. [12]
Поскольку скорости метаболических реакций зависят от концентрации активированных молекул и поскольку эти концентрации при каждом повышении температуры на 10 С увеличиваются в 2 - 3 раза, могут возникнуть серьезные проблемы, если колебания температуры тела не будут ограничены узкими пределами в несколько градусов. При снижении темпера-туры метаболические реакции могут настолько замедлиться, что необходимые для жизни процессы уже не будут протекать с достаточной скоростью. И наоборот, повышение температуры может привести к такому ускорению метаболических процессов, что снабжение организма достаточным количеством пищи и кислорода окажется невозможным. Кроме того, не для всех реакций величина Qio одинакова; поэтому изменение температуры будет вызывать дифференциальные эффекты, и метаболизм может быть выведен из равновесия. Так как эти проблемы существовали для живых систем с самого начала их эволюционной истории, мы могли бы предполагать, что в процессе эволюции выработались различные механизмы, позволяющие организмам контролировать интенсивность собственных жизненных процессов. И действительно, одно из эволюционных достижений, характерное для представителей большинства филогенетических линий, состоит в выработке способности поддерживать определенный уровень метаболической активности независимо от температуры окружающей среды. [13]
Показано, что ответная метаболическая реакция грибов определяется как структурой и свойствами фунгицидов, так и строением, характером обмена веществ грибов. Обращается внимание на зависимость фунгицидного эффекта от внеших условий и необходимость учета дополнительного фактора - свойств материала при подборе токсических веществ для их практического применения в народном хозяйстве. [14]
Однако не менее часто метаболические реакции подобного типа протекают как антимарковниковс-кие. Схема 29 показывает, что в этом случае из того же триена 2.177 возникают производные циклической системы бицикло [5.3.0] декана. Вещества с углеродным каркасом 2.288 относят к типу гваяна, а образующиеся из них в результате пинаколиновой перегруппировки структурные изомеры 2.289 именуют псевдогваянами. [15]
Страницы: 1 2 3 4