Регуляция - процесс
Cтраница 1
Регуляция процессов транспорта, как и регуляция процессов внутриклеточного метаболизма, осуществляется на двух уровнях: на уровне биосинтеза белковых посредников ( переносчиков) и на уровне функционирования готовых посредников. [1]
Регуляция процессов активного транспорта, обеспечивающего поступление подавляющего большинства необходимых прокариотам веществ, происходит на уровне синтеза переносчика и его функционирования. Биосинтез белковых компонентов многих транспортных систем регулируется по типу индукции. Глюкоза, транспортная система которой у большинства прокариот конститутивна, подавляет образование транспортных систем других Сахаров и ряда органических кислот путем катаболитной репрессии. Исключение составляют некоторые облигатно аэробные прокариоты, у которых транспорт органических кислот конститутивен, а индуцируемой является транспортная система глюкозы. Избыток субстрата в среде может репрессировать синтез соответствующей транспортной системы. Это особенно характерно для аминокислот. В этом случае регуляция транспорта координирована с регуляцией их последующего метаболизма. [2]
Регуляция че аболических процессов может осуществляться на разных уровнях постепенно возрастающей сложности. [3]
В регуляции процессов, протекающих в полевом сообществе ( агробиоценозе), большое внимание уделяют формированию смешанных посевов разных видов культурных растений. Экологические преимущества смешанных посевов перед чистыми известны давно. [4]
 |
Упрощенная схема основных элементов парасимпатической ( А и симпатической ( Б нервной системы. Сенсорные нейроны к автономной нервной системе не относятся. [5] |
Для регуляции процессов, не связанных с сокращением произвольной ( скелетной) мускулатуры, служат рефлексы, осуществляемые с помощью автономной ( вегетативной) нервной системы. [6]
В регуляции процессов образования и отдачи тепла в организме всегда участвует нервная система. Вообще организм приспособляется к окружающей среде в основном благодаря рефлексам, благодаря деятельности нервной системы. Последняя играет ведущую роль и в процессе закаливания. [7]
Непосредственное отношение к регуляции процессов, связанных с линькой, имеет функция щитовидной железы. Прежде считали, что естественная линька вызывается прямым действием гормона этой железы - тироксина. По современным данным, этот процесс более сложен: введение экзогенного тироксина не во всех случаях вызывает начало или ускорение линьки; тиреоэктомия или фармакологическая блокада щитовидной железы у птиц, начавших линьку, не вызывает ее прекращения. Предполагают, что тироксин необходим для регуляции подготовительных метаболических процессов, предшествующих началу смены оперения. Во время собственно линьки роль этого гормона менее значительна; линька может проходить и при низком уровне его секреции, на основе эндогенной программы. В этом смысле щитовидная железа сохраняет свою генеральную функцию контроля энергетического обмена. Участие же тироксина в регуляции конкретных сроков линьки определяется влиянием ряда факторов на активность щитовидной железы. В частности, функция этой железы в определенной степени регулируется и длительностью фотопериода При этом имеют место сезонные изменения типа реакции: во второй половине года секреция тироксина, по-видимому, усиливается короткими фотопериодами, а в первой половине года - длинными. Этого достаточно для стимуляции частичной линьки или для усиления уже начавшейся послебрачной линьки. [8]
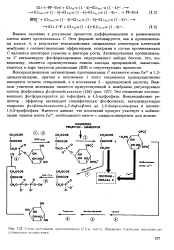 |
Схема регуляции протеинкиназы С ( текст. Жирными стрелками показаны ре-гуляторные воздействия. [9] |
Важное значение в регуляции процессов дифференцировки и размножения клеток имеет протеинкиназа С. Этот фермент активируется, как и протеинкина-зы класса А, в результате взаимодействия специальных рецепторов клеточной мембраны с соответствующими эффекторами, которыми в случае протеинкиназы С являются некоторые гормоны и факторы роста. Активированная протеинкиназа С катализирует фосфорилирование определенного набора белков, что, по-видимому, является промежуточным этапом каскада превращений, заканчивающегося в ядре запуском репликации ДНК и сопутствующих процессов. [10]
Селезенка активно участвует в регуляции процессов кроветворения, осуществляющихся костным мозгом. Она регулирует созревание и выход из костного мозга красных и белых кровяных телец, продукцию тромбоцитов мегакариоцитами, процесс денуклеации созревающих эритроцитов, продукцию лимфоцитов. Лучевого повреждения костного мозга не возникает, если перед общим облучением организма экранировать селезенку. [11]
 |
События в мембранах ( Витт. [12] |
С другой стороны, тонкая регуляция внутриклеточных процессов осуществляется на основе пространственного разделения органоидов клетки. Внутриклеточные мембраны служат для ком-партментации внутреннего содержимого клетки. [13]
Рассмотрим кратко вопрос о регуляции процессов дифференцировки клеток высших организмов. ДНК, присутствующая во всех соматических клетках, вероятнее всего, имеет одинаковую первичную структуру у данного организма и соответственно располагает информацией для синтеза любых или всех белков тела. Тем не менее клетки печени, например, синтезируют сывороточные белки, а клетки молочной железы - белки молока. Нет сомнения в том, что в дифференцированных клетках имеется весьма тонкий механизм контроля деятельности ДНК в разных тканях, обеспечивающий синтез многообразия белков. [14]
У животных важную роль в регуляции процессов роста и развития играют нервная и эндокринная системы. Наиболее сложен онтогенез многоклеточных животных, в котором выделяют следующие основные периоды ( этапы): от оплодотворения до выхода организма из яйцевых и зародышевых оболочек, или рождения, - эмбриональный; развитие и рост до половой зрелости - постэмбриональный; взрослое состояние, включая последующее старение организма. [15]
Страницы: 1 2 3 4