Регуляция - активность
Cтраница 3
Роль мотива ССААТ может быть достаточно важной в регуляции активности глобиновых генов, активируемых или репрессируемых на определенных стадиях развития. Репрессия синтеза - глобина плода у взрослого организма снимается в случае мутационного замещения одного нуклеотида в последовательности ССААТ. [31]
Роль мотива ССААТ может быть достаточно важной в регуляции активности глобиновых генов, активируемых или репрессируемых на определенных стадиях развития. Репрессия синтеза v-глобина плода у взрослого организма снимается в случае мутационного замещения одного нуклеотида в последовательности ССААТ. [32]
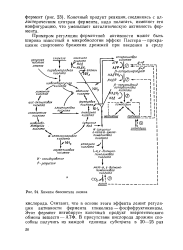 |
Химизм биосинтеза лизина. [33] |
Считают, что в основе этого эффекта лежит регуляция активности фермента гликолиза - фосфофруктокиназы. Этот фермент ингибирует конечный продукт энергетического обмена веществ - АТФ. [34]
Роль мотива ССААТ может быть достаточно важной в регуляции активности глобиновых генов, активируемых или репрессируемых на определенных стадиях развития. Репрессия синтеза v-глобина плода у взрослого организма снимается в случае мутационного замещения одного нуклеотида в последовательности ССААТ. [35]
Указанные положения составляют основу современной концепции адсорбционного механизма регуляции активности ферментов в клетке, в том числе и гексокиназы тканей млекопитающих. [36]
Роль моно - и поли - АДФ-рибозилирования белков в регуляции активности ферментов и модификации структуры хроматина. Поли-АДФ-рибозилирование белков приводит к нарушению их конформа-ции. В случае ядерных белков это влечет за собой изменение функционального состояния хроматина. Количество остатков АДФ-рибозы, присоединившихся к молекуле того или иного белка, зависит от активности как полимеразы, так и гидролазы. Следует учитывать также, что степень модифицируемости белков определяется не только активностью ферментной системы метаболизма поли - АДФ-рибозы, но и доступностью самих акцепторов для отдельных компонентов этой системы. В то же время, присоединяясь к ферментам, остатки АДФ-рибозы изменяют их каталитическую активность. Таким образом, система АДФ-рибозилирования белков может выполнять роль универсального и гибкого регулятора разнообразных биохимических процессов в клетке. [37]
Наконец, последний аспект фосфофруктокиназного катализа, повышающий точность регуляции активности ФФК в мышце, касается сродства между ферментом и субстратом. [38]
Конформационные изменения играют важную роль в функционировании ферментов и в регуляции активности ферментов. [39]
Пострадиационное нарушение этого процесса может быть следствием и повреждения системы регуляции активности генов, ответственных за синтез индивидуальных ферментов, которые катализируют отдельные этапы указанного процесса. Например, показано, что ионизирующие излучения вызывают дерепрессию ДНК-матрицы в печени и репрессию - в селезенке, тогда как скорость синтеза РНК на функционирующих цистронах не изменяется. Эти данные удовлетворительно коррелируют с экспериментально полученными результатами. [40]
Так как проблема компенсации сводится в данном случае к задаче регуляции активности ферментов, большая часть механизмов, эффективно регулирующих метаболические процессы в условиях постоянной температуры, потенциально могла бы быть пригодной и для компенсации температурных эффектов. [41]
Необходимо отказаться от использования в психологической теории алгоритмических и кибернетических принципов регуляции активности. Регуляция в алгоритмических и кибернетических машинах осуществляется по принципу предварительного установления порядка между специфическими инструкциями или материальными частями указанных машин соответственно. Однако этот принцип не позволяет адекватно понять антиэнтропийную сущность и развитие живых систем, а также приводит к пониманию их упорядоченности как априорного, заранее определенного факта. [42]
Нетрудно представить огромную роль ферментных систем или даже отдельных ферментов, нарушение регуляции активности и синтеза которых приводит к формированию или развитию патологического процесса. [43]
В белковой части фермента может находиться и аллостери-ческий центр, имеющий большое значение в регуляции ферментной активности. После присоединения к этому центру соответствующих веществ - эффекторов активность фермента изменяется. Конечные продукты ферментативных реакций обычно являются негативными эффекторами - присоединение их к ал-лостерическому центру фермента уменьшает его активность. Вещества, присоединение которых к аллостерическому центру молекулы фермента вызывают увеличение активности, называют позитивными эффекторами. [44]
Участвуют в поддержании и изменении структуры хромосом на разных стадиях клеточного цикла, в регуляции активности генов. ГИСТОХИМИЯ, раздел гистологии, изучающий хим. свойства тканей животных и человека. [45]
Страницы: 1 2 3 4