Гомологичная рекомбинация
Cтраница 2
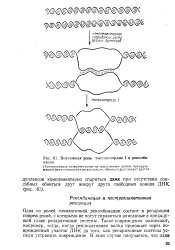 |
Возможная роль топоизомеразы I в рекомбинации. [16] |
Одна из ролей гомологичной рекомбинации состоит в репарации повреждений, с которыми. Такие повреждения возникают, например, тогда, когда репликативная вилка проезжает через поврежденный участок ДНК до того, как репаративные системы успели устранить повреждение. [17]
 |
Природные катаболические плазмиды. [18] |
Тесно сцепленные решшконы могут претерпевать гомологичную рекомбинацию, частота которой прямо пропорциональна степени структурной гомологии между плазмидами. [19]
 |
Генная конверсия ( о и один из возможных механизмов генной конверсии ( б. [20] |
У этого организма практически все виды гомологичной рекомбинации зависят от функционирования гена гее А. Оказалось, что RecA-белок ( молекулярная масса около 40 кД) является ДНК-зависимой АТРазой, способной заменять одну из цепей дуплекса ДНК на гомологичную. Для осуществления этой реакции RecA-белок покрывает одно-цепочечную ДНК и такой комплекс связывается с двуцепочечной ДНК. Процесс поиска является активным и требует гидролиза АТР. Показано, что дуплекс ДНК в комплексе с RecA-белком и одноцепочечной ДНК заметно раскручен ( вне зависимости от того, гомологична однонитевая ДНК дуплексу или нет); по всей вероятности, это облегчает поиск гомологии. Другими словами, RecA-белок активно проводит однонитевую миграцию ветви. [21]
 |
Генная конверсия ( а и один из возможных механизмов генной конверсии ( б. [22] |
У этого организма практически все виды гомологичной рекомбинации зависят от функционирования гена гее А. Оказалось, что RecA-белок ( молекулярная масса около 40 кД) является ДНК-зависимой АТРазой, способной заменять одну из цепей дуплекса ДНК на гомологичную. Процесс поиска является активным и требует гидролиза АТР. Показано, что дуплекс ДНК в комплексе с RecA-белком и одноцепочечной ДНК заметно раскручен ( вне зависимости от того, гомологична однонитевая ДНК дуплексу или нет); по всей вероятности, это облегчает поиск гомологии. [23]

У этого организма практически все виды гомологичной рекомбинации зависят от функционирования гена reck ( этот ген играет важную роль и в репарации повреждений ДНК; см. гл. Оказалось, что RecA-белок ( молекулярная масса около 40 кД) является ДНК-зависимой АТРазой, способной заменять одну из цепей дуплекса ДНК на гомологичную. Процесс поиска является активным и требует гидролиза АТР. Показано, что дуплекс ДНК в комплексе с RecA-белком и одноцепочечной ДНК заметно раскручен ( вне зависимости от того, гомологична однонитевая ДНК дуплексу или нет); по всей вероятности, это облегчает поиск гомологии. Другими словами, RecA-белок активно проводит однонитевую миграцию ветви. [25]
Ферментные системы клеток, с помощью которых осуществляется гомологичная рекомбинация, и тем более лежащие в основе рекомбинации молекулярные события еще недостаточно изучены. Основные сведения о процессах рекомбинации получены генетическими методами путем наблюдения за возникновением новых комбинаций наследуемых признаков в потомстве. [26]
Интеграция ( Integration) Встраивание чужеродной ДНК ( обычно с помощью гомологичной рекомбинации) в хромосому хозяйской клетки. [27]
Одинаковые IS-элементы и транспозоны, расположенные на разных репликонах, способны обеспечивать гомологичную рекомбинацию, приводящую к образованию коинтеграта. Плазмиды, способные встраиваться в хромосому бактерий и вырезаться оттуда, называют эписомами. Иногда вырезание эписомы может происходить не по той паре IS-элементов, по которой прошла интеграция. [29]
Мы видим, что по крайней мере у бактерий в общих чертах понятен механизм центрального события гомологичной рекомбинации поиска гомологии и образования полухиазмы. [30]
Страницы: 1 2 3 4