Ретровирус
Cтраница 2

Жизненный цикл ретровируса включает следующие стадии. [17]
Чтобы заразить рекомбинантным ретровирусом эмбриональные клетки, в культуральную емкость с инфицированными фиб-робластами, продуцирующими рекомбинантный вирус, помещают восьмиклеточную морулу ( группа бластомер, возникшая после равномерного дробления оплодотворенного яйца), освобожденную от яйцевой оболочки. Морула инфицируется и после достижения стадии бластоцисты ее вводят в матку псевдобеременной матери. Часть бластоцист может погибнуть, а часть нормально развивается и в соответствующие сроки трансформируется в трансгенное потомство, которое затем подвергается тщательному генетическому анализу и может использоваться для выведения трансгенных линий. [18]
Кроме того, ретровирусы вызывают очень слабый иммунитет. Более эффективно создание вакцин, содержащих фрагменты вирусного белка. [19]
Совершенно иначе функционируют ретровирусы. [20]
Известно, что многие ретровирусы могут переноситься между различными видами и даже классами позвоночных. Обнаружено также удивительное сходство между вирусами, выделенными у обезьян, с одной стороны, и ВИЧ, особенно ВИЧ-2 и ВЧТЛ-4 - с другой. [21]
ВИЧ относится к семейству ретровирусов. [22]
Тодаро выдвинули идею, что так называемые ретровирусы типа С могут встраиваться в генетический аппарат позвоночных и в составе их генома ( совокупность генов в наборе хромосом данного организма) передаваться от поколения к поколению. Они предположили, что эти ретровирусы содержат в своем геноме онкоген - который способен в определенных условиях активироваться и вызывать превращение нормальных клеток в опухолевые. [23]
Впервые подобный процесс был описан при исследовании ретровирусов, в геноме которых имеется ген, кодирующий обратную транскриптазу ( ревертазу) ( см. гл. В геноме млекопитающих, птиц, амфибий и насекомых обнаруживаются ретропо-зоны, представляющие собой внедрившиеся в геном ДНК-копии, синтезированные на разных типах клеточных РНК как на матрицах. Молекулярные механизмы ретропозиции не изучены, остается не установленным источник клеточной обратной тран-скриптазы. Не ясно, что служит затравкой для ревертазы; возможно, это шпилька на З - конце РНК, образующаяся в результате комплементарных взаимодействий. Как будет видно, структура ретропозонов позволяет с уверенностью говорить об участии обратной транскрипции в процессе их образования. Таким образом, наряду с переносом информации от ДНК к РНК осуществляется и обратный процесс - возвращение ее в геном в виде ретропозонов. У млекопитающих ретропозоны составляют более 10 % ДНК; следовательно, мощность встречного потока информации от РНК к ДНК может быть существенной, по крайней мере при оценке его во временном эволюционном масштабе. Различают разные типы ретропозоков. [24]
Впервые подобный процесс был описан при исследовании ретровирусов, в геноме которых имеется ген, кодирующий обратную транскриптазу ( ревертазу) ( см. гл. В геноме млекопитающих, птиц, амфибий и насекомых обнаруживаются ретропо-зоны, представляющие собой внедрившиеся в геном ДНК-копии, синтезированные на разных типах клеточных РНК как на матрицах. Молекулярные механизмы ретропозиции не изучены, остается не установленным источник клеточной обратной тран-скриптазы. Не ясно, что служит затравкой для ревертазы; возможно, это шпилька на З - конце РНК, образующаяся в результате комплементарных взаимодействий. Как будет видно, структура ретропозонов позволяет с уверенностью говорить об участии обратной транскрипции в процессе их образования. Таким образом, наряду с переносом информации от ДНК к РНК осуществляется и обратный процесс - возвращение ее в геном в виде ретропозонов. У млекопитающих ретропозоны составляют более 10 % ДНК; следовательно, мощность встречного потока информации от РНК к ДНК может быть существенной, по крайней мере при оценке его во временном эволюционном масштабе. Различают разные типы ретропозоков. [25]
Впервые подобный процесс был описан при исследовании ретровирусов, в геноме которых имеется ген, кодирующий обратную транскриптазу ( ревертазу) ( см. гл. В геноме млекопитающих, птиц, амфибий и насекомых обнаруживаются ретропо-зоны, представляющие собой внедрившиеся в геном ДНК-копии, синтезированные на разных типах клеточных РНК как на матрицах. Молекулярные механизмы ретропозиции не изучены, остается не установленным источник клеточной обратной тран-скриптазы. Не ясно, что служит затравкой для ревертазы; возможно, это шпилька на З - конце РНК, образующаяся в результате комплементарных взаимодействий. Как будет видно, структура ретропозонов позволяет с уверенностью говорить об участии обратной транскрипции в процессе их образования. Таким образом, наряду с переносом информации от ДНК к РНК осуществляется и обратный процесс - возвращение ее в геном в виде ретропозонов. У млекопитающих ретропозоны составляют более 10 % ДНК; следовательно, мощность встречного потока информации от РНК к ДНК может быть существенной, по крайней мере при оценке его во временном эволюционном масштабе. Различают разные типы ретропозоков. [26]
СПИД - первое эпидемическое заболевание человека, вызванное ретровирусами. [27]
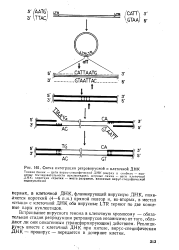 |
Схема интеграции ретровирусной и клеточной ДНК. [28] |
Встраивание вирусного генома в клеточную хромосому - обязательная стадия репродукции ретровирусов независимо от того, обладают ли они онкогенным ( трансформирующим) действием. Реплицируясь вместе с клеточной ДНК при митозе, вирус-специфическая ДНК-провирус - передается в дочерние клетки. [29]
Итак, ВИЧ, обладая общими свойствами, характерными для ретровирусов, имеет и некоторые существенные особенности. [30]
Страницы: 1 2 3 4