Опиатный рецептор
Cтраница 2
Опиаты обладают как анальгетическим, так и эйфорическим действием. Распределение опиатных рецепторов в головном мозге было исследовано с помощью радиоактивно меченных лигандов и флуоресцентно меченных антител против энкефалинов. В спинном мозге они непосредственно действуют на проводимость болевых ощущений. Исследование этого отдела ЦНС как раз дает прямые доказательства физиологической роли опиатных рецепторов. Они являются, вероятно, пресинаптическими соединениями и поэтому их следует искать в нервных окончаниях. [16]
Наиболее еажным является продукт О-деметилирования ТРМ - метаболит О-ТРМ. Он активен фармакологически, отличается большим сродством к опиатным рецепторам, чем сам ТРМ, и превышает его по анальгетической активности в 2 - 4 раза ( 15 16J, подобно тому, как продукт О-деметилирования КОДЕИНА, метаболит. [17]
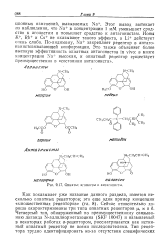 |
Опиаты. агонисты и антагонисты. [18] |
Как показывает уже название данного раздела, имеется несколько опнатных рецепторов; это еще один пример концепции множественных рецепторов ( гл. Четвертый тип, обнаруженный по преимущественному связыванию лиганда N-аллилнорметазоцина ( SKF 10047) и называемый в некоторых работах о-рецептором, рассматривается как истинный опиатный рецептор не всеми исследователями. [19]
В мозге были обнаружены специфические рецепторы этих лекарств, и фактически, как уже отмечалось, в настоящее время известно несколько типов опиатных рецепторов. Давно предполагалось, что должны существовать и эндогенные лиган-ды для этих рецепторов, но только в 1974 г. группами Костер-лица и Хьюза и Терениуса и Валстрема были открыты в экстрактах мозга пептиды, которые обладали свойствами, ожидаемыми для эндогенных опиатов. Пока еще неясно, являются ли эти пептиды истинными нейромедиаторами или просто нейро-модуляторами; здесь приведен краткий обзор имеющихся по этому вопросу данных. [20]

Пентапептид, включающий первые пять остатков с N-конца эндорфина, представляет собой так называемый энкефалин ( находящийся в мозгу), который связывается с опиатными рецепторами мозга и обладает очень высокой морфиноподобной активностью. Упомянутые выше открытия, сделанные в последние годы, ведут к созданию новых представлений о биохимических механизмах боли и наркомании. [22]
У гармана, его нсгидрированных ( гармина) и дегидрированных армалина) производных обнаруживается сродство к опиатным ре-шторам, но в микромолярных концентрациях ( см табл. 6.8) Пропето [447] сравнение влияния 0-карболинов на специфическое свя - 1вание 3Н - дигидроморфина ( / u - лиганда опиатных рецепторов) и 1-лейэнкефалина ( б-лиганда) с мембранами мозга. Найдено что Г50 / 3-карболинов, при которых наблюдается ингибирование связыва-3 Н - лейэнкефалина, намного меньше значений / С 0, ингибирующих гязывание ju - лигянда опиатных рецепторов т.е. 0-карболины являют-преимущественно б лигандами. Так как в присутствии Na 50 ммоль / л NaCl) сродство / 3-карболинов к рецеторам опиатов су-ественно уменьшается [447, 448], 0-карболины представляют собой [ гонисты энкефалинов и морфина. [23]
Как показано Анохиной и сотрудниками, алкоголизация самок во время беременности и лактации резко ( в 3 - 10 раз) повышает содержание циклических нуклеотидов в мозге и приводит к снижению содержания циклического аденозинмонофосфата в крови. Помимо этого, в крови подопытных крысят повышено содержание дофамина. Обнаружено также сходство в состоянии дофаминовых и опиатных рецепторов в ряде мозговых образований потомства алкоголизированных животных и взрослых животных, предпочитающих алкоголь. [24]
 |
Опиатный рецептор. модель двух состояний. Ионы Na способствуют переходу из конфигурации, связывающей агонист, в конфигурацию, связывающую антагонист. [25] |
Опиаты обладают как анальгетическим, так и эйфорическим действием. Распределение опиатных рецепторов в головном мозге было исследовано с помощью радиоактивно меченных лигандов и флуоресцентно меченных антител против энкефалинов. В спинном мозге они непосредственно действуют на проводимость болевых ощущений. Исследование этого отдела ЦНС как раз дает прямые доказательства физиологической роли опиатных рецепторов. Они являются, вероятно, пресинаптическими соединениями и поэтому их следует искать в нервных окончаниях. [26]
В 70 - х годах в головном мозге различных позвоночных животных были обнаружены специфические рецепторы морфина. Эти рецепторы сосредоточены на синаптических мембранах. Наиболее богата ими лимбическая система, от которой зависит эмоциональный ответ. Эти пептиды, обладающие способностью специфически связываться с опиатными рецепторами, получили название эндорфинов и энкефалинов. [27]
Морфин назван по имени бога сновидений. Этот алкалоид, выделенный из млечного сока незрелых коробочек мака, а также опиаты, его производные, нашли применение при характеристике рецепторов центральной нервной системы, имеющих отношение к болевой чувствительности. Поскольку такие рецепторы в норме являются центрами действия эндогенных физиологически активных соединений, открытие морфиновых рецепторов заставило предположить, что имеется и эндогенное вещество по своему действию. Это стало предпосылкой для выделения пептидов, подобных по действию опиатам, - эндорфина, ди-норфина и энкефалинов; они, вероятно, являются медиаторами; в мозге ( гл. Опиатный рецептор стереоспецифичен, он связывает только ( -) - изомеры опиатов ( рис. 9.17) и различает-агонисты и антагонисты. Сродство агонистов коррелирует с их. Достаточно странно, что опиатные рецеп - торы, помимо центральной нервной системы, найдены только в пищеварительном тракте и в семявыводящем протоке ( vag deferens) мыши. Здесь, однако, их функции не выяснены. [28]
Страницы: 1 2