Фосфодиэфирные связи
Cтраница 3
 |
Схема, иллюстрирующая экспериментальную проверку модели вторичной структуры РНК. [31] |
Известно большое число РНКаз, которые направленно гидролизуют фосфодиэфирные связи в одно-тяжевых участках РНК. [32]
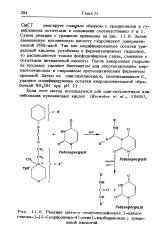 |
Реакция мето-п-толуолсульфоната 1-цикле - гексил-3 - [ 2 - ( морфолинил-4 - этил ] - карбодиимида с уриди. [33] |
Схема реакции с урацилом приведена на рис. 11.8. Затем замещенную нуклеиновую кислоту гидролизуют панкреатической РНК-азой. Так как модифицированные остатки ури - диловой кислоты устойчивы к ферментативному гидролизу, то расщепляются только фосфодиэфирные связи, смежные с остатками цитидиловой кислоты. После завершения гидролиза нуклеазу удаляют бентонитом или восстанавлением мер-каптоэтанолом и гидролизом протеолитическим ферментом проназой. [34]
В глицеринтейхоевых кислотах глицериновые единицы соединяются через фосфодиэфирные группы в 1 - м и 3 - м положениях. Но есть глицеринтейхоевые кислоты, имеющие связь между первым и вторым положениями, а также такие, где фосфодиэфирные связи соединяют не только полиоловые, но и углеводные единицы. [35]
После того как было установлено, что рибонуклеиновые кислоты состоят в основном из четырех мононуклеотидных единиц, в течение многих лет отсутствовали точные сведения относительно характера межнуклеотидных связей и поэтому было высказано множе - ство предположений. Многие предполагаемые структуры включали пирофосфатные, полифосфорные, эфирные и фосфоамидные связи, но относительно простая тетрануклеотидная структура, предложенная Левиным [65, 66] и содержавшая фосфодиэфирные связи между углеводными компонентами нуклеозидов, лучше всего, как позже было выяснено, соответствовала действительности. Хотя в настоящее время тетрануклеотидная теория строения нуклеиновых кислот полностью оставлена, уместно, быть может, упомянуть, что эта теория была в свое время значительно точнее тринуклеотидной теории [67, 68], с которой она находилась в оппозиции, и что, как писал сам Левин, с другой стороны, нужно иметь в виду, что истинный молекулярный вес нуклеиновых кислот до сих пор еще неизвестен. [36]
Фрагментация нуклеотидной цепи при анализе последовательности осуществляется с помощью ферментов, называемых нуклеазами. Известно значительное число таких ферментов. Одни из них расщепляют фосфодиэфирные связи внутри пол и нуклеотидной цепи ( эидодезоксирибонуклеазы), другие гидролизуют цепь начиная с 5 - или З - конца - они называются 5 -или 3 -экзодезоксирибонуклеазами. Эидоиуклеазы различаются по специфичности - расщепляют определенные последовательности или не предъявляют к последовательности никаких требований. Существуют ферменты, гидролизующие только одноцепочеч-ные ДНК или двухцепочечные ДНК, или те и другие. Наиболее важным классом эидодезоксирибонуклеаз являются рестрикцион-ные эидоиуклеазы. [37]
Химический гидролиз ДНК почти не применяют из-за ослож-ния его побочными процессами. Более предпочтителен фермен-тивный гидролиз под действием нуклеаз. Обычно для этой цели пользуют змеиный яд, в котором содержатся ферменты, рас-епляющие фосфодиэфирные связи. [38]
Рибонуклеозидтрифосфаты и дезоксирибонуклеозидтрифосфаты не являются субстратами фермента. Фермент не нуждается в матрице, однако для синтеза необходима затравочная цепь РНК ( НМФ) п со свободной З - гидроксильной группой, к которой присоединяются остатки моно-нуклеотидов. Образовавшаяся полимерная молекула РНК не имеет заданной специфической последовательности мононуклеотидов, но содержит 3 - 5 фосфодиэфирные связи, легко разрываемые рибонуклеазой. Относительно биологической роли этого фермента у бактерий предполагают, что он катализирует, скорее всего, обратную реакцию-расщепление мРНК с образованием нуклеозиддифосфатов. [39]
Дизамещенные фосфаты гидролизуются фер ( ментами фосфо-диэстеразами. Некоторые из них неспецифичны, но большинство участвует в шдролизе нуклеиновых кислот и называется нуклеаза-ми. Существуют рибонуклеазы и дезоксирибонуклеазы ( гл. Экзонуклеазы атакуют фосфодиэфирные связи последовательно, начиная с одного конца цепи нуклеиновой кислоты, в то время как эндонуклеазы атакуют также и связи внутри цепи. [40]
 |
Возможная схема действия ДНК-лв-гаэ. Цифры в кружках / - 6 обозначают последовательность действия фермента. [41] |
ДНК-лигазы - ферменты, катализирующие репарацию одноцепочечного разрыва, который обычно возникает под влиянием эндонуклеаз. Поэтому ДНК-лигазы называют еще и сшивающими ферментами. Лигазы обнаружены в самых различных клетках, в том числе и в клетках, зараженных вирусами. Под влиянием этих ферментов образуются фосфодиэфирные связи между свободным 5 -фосфатным концом олиго - или полинуклеотида и З - гидроксильной группой соседнего олиго - или полинуклеотида. Скорее всего реакция протекает с обязательным образованием промежуточного комплекса лигазы с АМФ. Например, в клетках кишечной палочки такой комплекс образуется при взаимодействии лигазы с НАД, а в клетках млекопитающих такой комплекс образуется при взаимодействии лигазы с АТФ. [42]
 |
Добавление липких концов к фрагменту ДНК, имеющему тупые концы, перед встраиванием ДНК в вектор с целью создания рекомбинантной молекулы ДНК. [43] |
Если для выделения донорной ДНК ( например, методом дробовика) использовали рестриктазу, той же самой рестриктазой должна быть обработана плазмидная ДНК. Рестрикци-онные фрагменты донорной ДНК, среди которых есть и фрагмент с нужным геном, смешивают затем с плазмидной ДНК. При смешивании они соединяются липкими концами. Вначале соединение происходит за счет водородных связей, но после добавления фермента, называемого ДНК-лигазой, образуются фосфодиэфирные связи. [44]
Однако если проводить гидразинолиз при достаточно низких значениях рН и низкой температуре, то удается выделить довольно высокомолекулярную апиримидиновую ДНК. В продуктах реакции не было обнаружено пири-мидинов, а константа седиментации полученной апиримидиновой ДНК мало отличалась от константы седиментации исходной ДНК. Продукт модификации седиментировал узкой, неразмытой зоной, что свидетельствует о малой деградации полинуклеотидной цепи. Из полученной апиримидиновой кислоты действием альдегидов ( например, бензальдегида) могут быть удалены остатки гидразина, и в конечном счете на месте пиримидинового нуклеотида в полинуклеотидной цепи оказывается негликозилированный рибозиль-ный остаток, фосфодиэфирные связи которого очень лабильны и могут быть расщеплены в мягких условиях ( см. стр. Апири-мидиновые нуклеиновые кислоты вследствие этого легко могут быть расщеплены до олигопуриновых нуклеотидов, и это широко применяется при анализе распределения пуринов в ДНК и РНК ( см. стр. Следует отметить, что образующиеся под действием гидразина апиримидиновые ДНК при обработке дифениламином и муравьиной кислотой ( см. стр. Образующийся при этом 4 - э / сзо - М - ами-ноцитозин может оказаться значительно более устойчивым к расщеплению. Тем не менее, расхождения экспериментальных значений с предсказываемыми теоретически для полностью апиридино-вой ДНК не очень велики. [45]
Страницы: 1 2 3 4