Взаимодействие - ген
Cтраница 1
Аллельное и неаллельное взаимодействие генов при самоопылении в значительной мере утратит свое действие на признак, так, как в результате гомозиготизации распадутся их эффективные комбинации, составные элементы которых распределятся по разным линиям. Правда, некоторые эффективные комбинации неаллельного взаимодействия генов ( эпистаз) могут сохраниться, но их эффект будет в данном случае аналогичен эффекту аддитивных генов и будет суммироваться с последними. [1]
Другой поразительный пример взаимодействия генов обнаружен у Lathyrus odoratus; было обнаружено, что мутация в локусе Ег контролирующем у этого растения гидроксили-рование антоцианидина, по-видимому, изменяет специфичность гликозидсинтезирующих ферментов. Должно быть, фермент, контролирующий синтез 3-латирозидо - 7-рамнозида флавонола у доминантных форм, у рецессивных мутантов утрачивает часть своей специфичности. [2]
Скорее всего, это можно объяснить особенностями взаимодействия генов с тканевыми структурами, в области которых они функционируют. К таким особенностям может, например, относиться разница во времени включения ( и выключения) тех или иных генов, последовательно кодирующих элементарные проявления иерархически устроенного фенотипического признака, в результате чего суммарный эффект генетических влияний оказывается различным на правой и левой половине тела. [3]
Чем существенно отличается взаимодействие ще лочных металлов с водородом от взаимодействия гало генов с водородом. Приведите пример той и другой ре акции и разберите эти реакции с точки зрения окисления - восстановления. [4]
Классической моделью для изучения плейотропии и биохимических процессов, лежащих в основе взаимодействия генов, является меланогенез. [5]
Генетик занимается проблемами генного взаимодействия в ограниченных масштабах, тогда как селекционер повсеместно сталкивается с результатами, обусловленными генным взаимодействием, и применяет их в своей деятельности, и в этом его сила, а не слабость. Ведь взаимодействие генов имеет дискретную почву с большим числом вариантов, далеко еще не охваченных генетическим изучением. [6]
 |
Эпистпаз у томата. [7] |
Когда два гена определяют одну и ту же биохимическую реакцию, соотношение фенотипов дает основание предполагать действие генов. Такое явление взаимодействия генов называется эпистазом. Различные типы взаимодействия генов создают различные соотношения F2 как результат слияния различных генотипических классов, имеющих сходные фенотипы. [8]
 |
Схема сочетания цепей действия генов ( по А. Кшпу. [9] |
Для построения белковых гранул, несомненно, нужна цепь действия генов, из которой при мутации wa - wa вырывается одно звено. Здесь хорошо выявлен модельный характер взаимодействия генов при построении клеточных структур. [10]
Аллельное и неаллельное взаимодействие генов при самоопылении в значительной мере утратит свое действие на признак, так, как в результате гомозиготизации распадутся их эффективные комбинации, составные элементы которых распределятся по разным линиям. Правда, некоторые эффективные комбинации неаллельного взаимодействия генов ( эпистаз) могут сохраниться, но их эффект будет в данном случае аналогичен эффекту аддитивных генов и будет суммироваться с последними. [11]
Наш экскурс в вопросы наследуемости кукурузы, кур и свойства футбольных полей иметь ширину был необходим для прояснения точного и очень ограниченного смысла понятия наследуемости, чтобы, когда мы услышим, что 7Q на 80 % наследуется, у нас было ясное понимание смысла этой фразы - независимо от того, правильна она или нет. Она не имеет ничего общего с действительным взаимодействием генов и среды, так же как свойство иметь ширину не имеет ничего общего с действительной длиной и шириной или с изменчивостью площадей или периметров. Наследуемость не связана с той ролью, которую играет наследственность при противопоставлении ее внешней среде. [12]
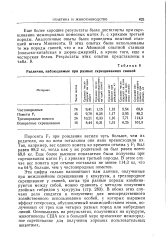 |
Различия, наблюдаемые при разных скрещиваниях свиней. [13] |
Эти цифры сильно напоминают нам данные, полученные при межлинейном скрещивании у кукурузы, а трехпородное скрещивание у свиней, когда три разные породы комбинируются между собой, можно сравнить с методом получения двойных гибридов у кукурузы ( стр. В обоих этих случаях дело сводится к получению для непосредственного хозяйственного использования особей, у которых выражен эффект гетерозиса, обусловленный взаимодействием генов, полученных от трех или четырех разных пород или инбредных линий. [14]
Необходимое приложение мутагенного начала в естественном отборе является верным и сильным постулатом, который выиграет от развертывания мутагенных опытов в некоторой микроэволюционной сфере. Здесь особенно важно установление времени, которое затрачивается на то, чтобы новый индуцированный признак был принят отбором; того, сколько индуцированных признаков принимается им одновременно и какой вес взаимодействия генов одного или разных организмов в сложных преобразованиях. [15]
Страницы: 1 2