Пигментная система
Cтраница 2
Аналогичная картина происходит и в пигментной системе II, у которой роль акцептора электрона выполняет пластохинон, передавая полученный электрон по электрон-транспортной цепи цитохрому Ь, цитохрому / и, наконец, пла-стоцианину. [16]
Пигментная система фоторецепторов сходна с пигментной системой фотосинтеза в том отношении, что в обоих случаях у ограниченного числа пигментов встречаются самые разные спектральные чувствительности, что объясняется разнообразием их взаимодействий с белками. [17]
Существенные аргументы в пользу функционирования двух пигментных систем следуют из изучения флуоресценции. [18]
Различные внешние факторы могут влиять на состав пигментной системы. Следует различать два вида адаптации - филогенетическую и онтогенетическую. С одной стороны, состав пигментной системы класса или вида связан с условиями, в которых он обычно живет. [19]
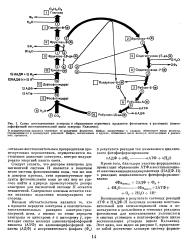 |
Схема восстановления углерода и образования первичных продуктов фотосинтеза в растениях ( пенто-зофосфатпый восстановительный цикл углерода Кальвина. [20] |
Следует сказать, что донором электрона для пигментной системы II является в конечном итоге система фотоокисления воды, так же как и донором протона, хотя промежуточные продукты фотоокисления воды до сих пор не удалось найти и природа промежуточного донора электрона для пигментной системы II остается неизвестной. Совершенно неясным остается также механизм выделения молекулярного кислорода. [21]
 |
Упрощенная схема цикла Кальвина. [22] |
Эта реакция реализуется в полной системе с участием пигментных систем и цепи переноса электронов. [23]
В табл. 22 показано, как эти изменения пигментной системы влияют на поглотительную способность растений в различных спектральных областях. Однако эта таблица относится к освещению светом с равномерным распределением интенсивности по спектру. [24]
В этом случае допускается, что оптические свойства пигментной системы могут быть описаны двумя константами и р, называемыми соответственно коэффициентом поглощения и коэффициентом рассеяния. [25]
По данным Мора, у проростков Sinapis alba обе пигментные системы действуют синергистически, вызывая образование антоциана, появление волосков на гипокотиле и стимулируя новообразование и рост зачатков листьев; угнетение роста гипокотиля обусловлено только активностью высокоэнергетической системы. [26]
С другой стороны, многие растения способны к индивидуальным изменениям пигментной системы. Одноклеточные зеленые водоросли, например Chlorella, становятся темнозелеными, если выращиваются на слабом свету, и светлозелеными, если они выращены на сильном свету. Красные Florideae становятся оливково-бурыми или даже зелеными, если их выставить на прямой солнечный свет. В некоторых случаях эти онтогенетические приспособления медленны и устойчивы, в других-они сравнительно быстрые и обратимые. Наиболее наглядным примером могут служить некоторые сине-зеленые водоросли, которые, как хамелеон, меняют окраску, реагируя на изменения в окраске света. [27]
Таким образом, вопрос о наличии, помимо фитохрома, высокоэнергетической пигментной системы и о взаимодействии этих двух систем остается открытым и требует дальнейших исследований. [28]
Следовательно, по крайней мере при комнатной температуре, испускание света пигментной системой I очень мало. Очевидно, кванты, не использованные для фотохимической реакции в этой системе, теряются путем безызлучатель-ных переходов. Возможно, эта потеря происходит в длинноволновой фракции хлорофилла С700, которая является предпоследним участником цепи ( фиг. Этот пигмент флуоресцирует только при низкой температуре и ответствен за полосу испускания при 700 ммк. [29]
Считают, что в переносе электронов воды к НАДФ участвуют последовательно две пигментные системы, которые содержат различные формы хлорофилла а, отличающиеся максимумами поглощения в длинноволновой части спектра. В первую систему входят также ка-ротиноиды, а во вторую - хлорофилл b и ряд других вспомогательных пигментов. [30]
Страницы: 1 2 3 4