Скорость - дыхание
Cтраница 3
Одним из основных параметров, характеризующих обмен выделенных митохондрий, является их способность к поглощению кислорода и зависимость скорости дыхания от присутствия акцепторной системы ( АДФ Фн) ( см. также с. В связи с этим для изучения метаболизма митохондрий необходимо иметь метод, позволяющий точно измерить поглощение кислорода при окислении митохондриями тех или иных субстратов. [31]
На рис. 7 представлены записи потенциалов действия диа-фрагмального нерва у кошек вместе с показаниями артериального давления и изменения скорости дыхания. На рис. 8 дана активность диафрагмального нерва в контрольных условиях. [32]
Соответствующая прямая должна пересекать ось абсцисс в точке Г, а ось ординат - при отрицательном значении, равном скорости темнового дыхания. Подобная зависимость была получена Га-астрой для репы, хотя, как видно из фиг. Если концентрация СО2 имеет тенденцию буферить в точке Г ( стр. Это может быть вызвано, например, влиянием концентрации СО2 на скорость дыхания на свету. [33]
Эти результаты, полученные Брауном в 1953 г., как будто бы ясно показывают, что обычная практика внесения поправки на скорость темнового дыхания правомочна. Однако, поскольку многие из промежуточных химических продуктов фотосинтеза и дыхания являются общими для обоих процессов, трудно допустить, что свет вообще не оказывает влияния на дыхание. Естественно было бы ожидать, что прежде чем субстраты дыхания деградируют до СОа и воды, часть их будет использована в синтетических процессах. [34]
Очевидно, в самой структуре митохондрий образование и связывание ионов Н и ОН - может служить важнейшими рычагами, регулирующими соотношение скоростей дыхания и фосфорилирования, и быть важнейшими элементами процессов активного переноса ионов. [35]
Шумы оказывают глубокое раздражающее влияние на весь организм человека: замедляют психические реакции, вызывают раздражительность, ускоряют процесс утомления, изменяют скорость дыхания и пульса, нарушают обмен веществ. [36]
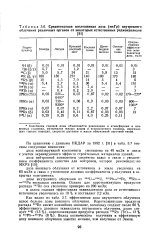 |
Среднегодовая поглощенная доза ( мкГр внутреннего облучения различных органов от некоторых естественных радионуклидов. [37] |
Колебания годовой дозы объясняются различиями в атмосферных и почвенных условиях, материалах жилыл домов и используемого топлива, окружающих горных породах, скорости дыхания и массе облучаемой легочной ткани. [38]
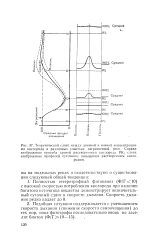
Полностью гетеротрофный фитоценоз ( Ф / Г10) с высокой скоростью потребления кислорода при наличии богатого источника вещества демонстрирует незначительный суточный сдвиг в скорости дыхания. [40]
Если считать правильными значения скорости фотодыхания, полученные описанным выше ( не вполне прямым) методом, то оказывается, что при концентрации кислорода ниже 10 % скорость дыхания, измеряемая по выделению COz, на свету ниже, чем в темноте ( фиг. Это было показано Брауном и Вейсом ( стр. Озбуном с сотрудниками для листьев фасоли при низких концентрациях кислорода, хотя в то же время указанные авторы нашли, что поглощение О2 на ярком свету увеличивается. [41]
При введении небольших доз брадикинина ( 1 - 8 мкг / кг внутривенно) анестезированным уретаном морским свинкам и кроликам наблюдается довольно существенное, но непродолжительное увеличение скорости дыхания. Стимулирующее действие брадикинина продолжается 4 - 8 мин; в этот период частота и объем дыхания увеличиваются в два-три раза. [42]
Помимо размеров как на степень, так и на преимущественное место задержки оказывают влияние плотность частиц, их форма, гигроскопичность, электрический заряд, а также частота и скорость дыхания и емкость легких. Следует иметь в виду, что по мере увеличения количества задержанных пылевых частиц в верхних дыхательных путях возможность их дальнейшей задержки уменьшается. [43]
СОз в зависимости от интенсивности света при 25 С и при концентрации СО2 0 03 %; II - теоретический график для истинного фотосинтеза, построенный в предположении, что скорость светового дыхания равнялась скорости тем-нового вплоть до компенсационного пункта, а затем линейно увеличивалась с увеличением интенсивности света. [44]
Средние наклоны в точке Г для верхней и нижней частей кривых регрессии значительно круче, чем наклон прямой, проходящей через точку Г и точку на оси ординат, соответствующую скорости темнового дыхания ( - 1); а регрессии ниже точки Г имеют заметные ( положительные) квадратичные коэффициенты и потому искривлены. Эти данные опровергают тривиальную гипотезу о простом балансе между скоростью фотосинтеза, пропорциональной, как полагают, наружной концентрации СОг, и постоянной скоростью дыхания ( фиг. [45]
Страницы: 1 2 3 4