Скорость - дезаминирование
Cтраница 2
Дезаминирование фотогидратов ци-тозиновых производных протекает медленнее, чем дегидратация. Скорость дезаминирования цитидин-3 - фосфата зависит от рН среды. [16]
Скорость дезаминирования быстро увеличивается при понижении рН; удивительно, что протонированная форма нуклеозида подвергается атаке электрофильного реагента быстрее, чем нейтральная форма. При понижении температуры реакции различие в скорости дезаминирования разных нуклеозидов несколько увеличивается. [17]
Обе реакции подчиняются закономерностям реакций первого порядка, и константы скорости реакций дегидратации и дезаминирования при рН 5 и 25 С равны соответственно 1 6 - 10 - 2 и 4 - 1 ( Н мин-1. При других значениях рН и температуры скорость дегидратации по-прежнему значительно выше скорости дезаминирования. Скорость замещения аминогруппы резко возрастает в присутствии различных нуклеофильных реагентов. [18]
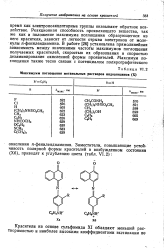 |
Максимумы поглощения метанольных растворов индоанилинов ( X. [19] |
Реакционная способность проявляющего вещества, так же как и положение максимума поглощения образующегося из него красителя, зависит от легкости отрыва электронов от молекулы я-фенилендиамина. В работе [26] установлена прямолинейная зависимость между значениями частоты максимумов поглощения получаемых красителей, скоростью их образования и скоростью дезаминирования окисленной формы проявителей. [20]
Непригодность этих аминокислот для обеспечения роста объясняется, вероятно, тем, что скорость их инверсии относительно невелика. Поскольку соответствующие лейцину и изолейцину а-кетокислоты легко используются для роста ( табл. 16), можно заключить, что лимитирующим фактором является скорость дезаминирования этих аминокислот. То, что D-лейнин действительно подвергается инверсии в организме, подтверждено исследованиями, в которых крысам вводили меченный дейтерием D-лейцин. [21]
Страницы: 1 2