Полиплоидный вид
Cтраница 1
Знание встречаемости полиплоидных видов и форм в пределах популяций имеет значение не только для теории эволюции, видообразЪвания и цитотаксономии, но и для интродукции полезных дикорастущих растений в культуру с целью использования их в фармакологии, парфюмерной и пищевой промышленности. Оно может быть критерием, определяющим направление поисков наиболее продуктивных видов и форм. [1]
Однако у полиплоидных видов моносомики жизнеспособны, поскольку утрата одной хромосомы сказывается у них гораздо слабее. Например, у обычного табака Nicotiana taba-cum тетраплоидного вида с 4 - 12 48 хромосомами известна серия различных моносомиков с 47 хромосомами. Кроме того, моносомия вызывает характерные изменения внешнего вида ( фиг. В профазе и метафазе мейоза единственная моносомная хромосома ведет себя как унивалент. [2]
Рассмотрен вопрос о происхождении полиплоидных видов и их цитогенети-ческих особенностей, определяющих способ использования этих видов в селекции. [3]
Приведен обзор работ по изучению геномного состава полиплоидных видов картофеля, относящихся к разным таксономическим сериям. Приведены их геномные формулы. Южноамериканские полиплоидные виды содержат серии генома А, североамериканские - серии геномов А и В. [4]
В отличие от диплоидов, геномная дифференциация у полиплоидных видов, особенно у гексаллоидных видов серии Demissa и тетраплоидных видов серии Longipedicellata, изучена значительно полнее. Имеющиеся данные показывают, что в противоположность диплоидам все полиплоидные виды картофеля имеют строго дифференцированные геномы. [5]
В этой связи изучение феномена ядрышкового доминирования, широко распространенного в растительном царстве при формировании полиплоидных видов растений, представляет значительный интерес. Для лучшего понимания процессов интеграции разнокачественных геномов при образовании полиплоидных форм необходима информация о тонкой структурно-функциональной организации и основных принципах эволюции генов рибосомных РНК. Твердая и мягкая пшеницы представляют собой, как известно, амфидиплоидную и аллополиплоид-ную формы, характеризующиеся дифференциальной экспрессией генов рРНК, расположенных на разных хромосомах, за счет весьма сильно выраженного эффекта ядрышкового доминирования, природа которого до конца не ясна. Поскольку промоторная область, да и субповторы, вероятно, вносят решающий вклад в уровень транскрипции генов рРНК, то изучение их организации, определение участков, непосредственно взаимодействующих с ядерными белками, и влияние отдельных нуклео-тидов могут способствовать пониманию механизмов ядрышковой доминантности. Для выяснения особенностей и условий возникновения феномена ядрышкового доминирования при интеграции разнокачественных геномов в процессе формирования полиплоидных видов пшениц необходимо изучение этих элементов межгенного спейсера рДНК у разных видов злаков трибы Triticeae, являющихся донорами пшеничных геномов или их ближайшими родственниками. [6]
Семейство крестоцветных ( Cruciferae), которые в основном числе своих представителей характеризуются мелкими хромосомами, также имеет большое количество полиплоидных видов и форм, среди которых 32 % выше гексаплоидного уровня. Среди изученных нами 24 среднеазиатских представителей семейства губоцветных, распространенных в Туркменской ССР, 11 видов были полиплоидными. Этим подтверждается значительная роль полиплоидии в видообразовании в пределах данного семейства, причем некоторые из них несомненно амфидиплоидного происхождения. [7]
Помимо пшеницы, имеются еще другие аллополиплоидные виды культурных растений, для которых известны исходные родительские виды; в таких случаях также оказалось возможным получить новые синтетические формы этих полиплоидных видов. Так была получена синтетическая форма обыкновенного табака Nicotiana tabacum, который представляет собой аллотетраплоид с 48 хромосомами, возникший путем объединения хромосом двух диплоидных видов: N. [8]
Однако в таких родах, как Triticum, Hordeum, у которых хромосомы относительно крупные ( 5 - 10 мк), уровень полиплоидиии не выше гексаплоидного, а в роде Secale, где хромосомы еще крупнее ( до 11 - 13 мк), полиплоидных видов совсем не обнаружено. [9]
Мы сопоставили встречаемость полиплоидов с наличием хромосом определенного размера у представителей семейств, разделив их условно на обладателей мелких ( 1 - 4 мк) и крупных ( 5 - 10, 15 мк) хромосом. Количество полиплоидных видов среди изученных цитологически достигает 32 6 % при наличии 16 % полиплоидных форм в пределах самих видов. [10]
Так называемые полигаплоиды ( особи, содержащие половинное число хромосом и развившиеся из полиплоидных видов) содержат несколько геномов. Термин гаплоид используется также для того, чтобы подчеркнуть, что гаметы ( у диплоидных видов) содержат только по одной хромосоме каждого типа. Таким образом, гаметы гаплоидны, а соматические клетки диплоидны. [11]
ДНК к диплоидному, а тетраплоидный вид имел приблизительно половину количества ДНК октоплоидного вида. Ротфельс и Хаймбургер [77] считают, что, хотя полиплоиды рода Drosera имеют более мелкие хромосомы, чем диплоиды, это скорее говорит не о необходимости филогенетической редукции, а о том, что диплоиды с хромосомами малых размеров дали начало полиплоидным видам. Пробст [78] также показал, что в ядрах меристемных клеток корешков, окрашенных по Фельгену, у вида Leucanthemum vulgare в полиплоидных формах ( а именно у октоплоидной с 2п72) содержание ДНК меньше, чем ожидалось теоретически. Рассматривая изображение метафазных пластинок ряда видов злаковых в работе Н.П. Авдулова [79], можно найти примеры изменения размеров хромосом в сторону их уменьшения по мере увеличения уровня полиплоидии, хотя сам автор этого не отмечает. Например, в роде Andropogon у видов, образующих полиплоидный ряд: A. [12]
Более того, как предполагают японские ученые С. Отсюда следует, что самый низкий современный уровень полиплоидии в подсемействе - гексаплоидный. Интересно, что процент полиплоидных видов в этом подсемействе значительно выше в тропических и южных умеренных зонах по сравнению с северными умеренными. Но главным центром видообразования этой группы папоротников являются тропики. [13]
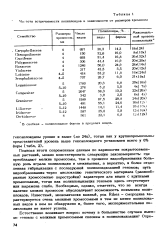 |
Частота встречаемости полиплоидов в зависимости от размеров хромосом. [14] |
Подводя итоги современным данным по кариологии покрытосеменных растений, можно констатировать следующую закономерность: где преобладают мелкие хромосомы, там в процессе видообразования большую роль играла полиплоидия и межвидовая, а вероятно, и более отдаленная гибридизация с последующей полиплоидизацией геномов; путь видообразования через дислокацию генетического материала ( разнообразные хромосомные перестройки) характерен для видов с крупными хромосомами, тогда как полиплоидизация геномов здесь отсутствует или выражена слабо. Необходимо, однако, отметить, что не всегда наличие мелких хромосом обусловливает возможность появления полиплоидов. Известный, экономически важный, род Ribes - смородина характеризуется очень мелкими хромосомами и тем не менее полиплоидных видов в нем не обнаружено и, более того, экспериментальная полиплоидия не имеет успеха. [15]
Страницы: 1 2