Комплементарное спаривание
Cтраница 1
Комплементарное спаривание PHKI с РНКП так меняет вторичную структуру последней, что она не может гибридизоваться с областью инициации синтеза ДНК - Интересно, что участок регуляторного воздействия, спаривания РНКН с PHKI отстоит на несколько сотен нуклеотидов от того места, где происходит регулируемое событие - гибридизация РНК с ДНК - Как и в других аналогичных случаях ( например, при регуляции активности генов биосинтеза аминокислот, см. гл. VII), дело здесь в том, что РНКП может в принципе образовать два различных, причем взаимоисключающих варианта вторичной структуры. Лишь один из вариантов способен гибридизоваться с ориджином, а спаривание с PHKI стабилизирует другой вариант и тем самым исключает репликацию. [1]
Комплементарное спаривание PHKI с РНКП так меняет вторичную структуру последней, что она не может гибридизоваться с областью инициации синтеза ДНК. Интересно, что участок регуляторного воздействия, спаривания РНКП с PHKI отстоит на несколько сотен нуклеотидов от того места, где происходит регулируемое событие - гибридизация РНК с ДНК - Как и в других аналогичных случаях ( например, при регуляции активности генов биосинтеза аминокислот, см. гл. VII), дело здесь в том, что РНКП может в принципе образовать два различных, причем взаимоисключающих варианта вторичной структуры. Лишь один из вариантов способен гибридизоваться с ориджином, а спаривание с PHKI стабилизирует другой вариант и тем самым исключает репликацию. [2]
Комплементарное спаривание PHKI с РНКП так меняет вторичную структуру последней, что она не может гибридизоваться с областью инициации синтеза ДНК - Интересно, что участок регуляторного воздействия, спаривания РНКП с PHKI отстоит на несколько сотен нуклеотидов от того места, где происходит регулируемое событие - гибридизация РНК с ДНК - Как и в других аналогичных случаях ( например, при регуляции активности генов биосинтеза аминокислот, см. гл. VII), дело здесь в том, что РНКП может в принципе образовать два различных, причем взаимоисключающих варианта вторичной структуры. Лишь один из вариантов способен гибридизоваться с ориджином, а спаривание с PHKI стабилизирует другой вариант и тем самым исключает репликацию. [3]
Ввиду существования комплементарного спаривания оснований некоторые динуклсотидныс последовательности должны встречаться одинаково часто. Равенство или неравенство частот данных динуклсо-тидных последовательностей зависит от того, как ориентированы цепи: параллельно или антипараллельно. [4]
В процессе комплементарного спаривания оснований информация, заключенная в последовательности кодогенов ДНК, переснимается, копируется в молекулу мРНК и в этом виде ( негатив) переносится от своего первоисточника к месту белкового синтеза, ибо, как нам уже известно, белковый синтез происходит вдали от источника информации - ДНК, ни и не непосредственно на мРНК - это было бы слишком просто. Где же поме-щается фабрика белка, где происходит расшифровка и использование переносимой РНК-посредником информации, ее перевод в последовательность аминокислот. [5]
Считается, что комплементарное спаривание З - концевого участка 16S РНК с прединициирующим полипуриновым блоком мРНК прямо участвует в ассоциации свободной рибосомной 30S субчастицы с матричным полинуклеотидом. [6]
В результате принципа комплементарного спаривания оснований молекула РНК точно отражает последовательность оснований в молекуле ДНК. Так, мРНК додержит остатки аденина там, где ДНК содержит тимин, остатки цитозина там, где ДНК содержит гуанин, гуанин там, где ДНК содержит цитозин, и остатки ураци-ла там, где ДНК содержит аденин. Но как это трансформируется в специфическую последовательность аминокислотных остатков молекулы белка. Это и есть самая интересная часть загадки. [7]
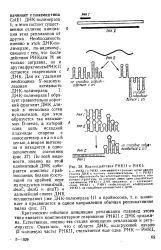
Критическим событием инициации репликации плазмид этого типа является комплементарное спаривание РНКИ с ДНК ориджи-на. Естественно, именно эта стадия инициации репликации и регулируется. [9]
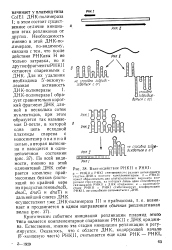 |
Взаимодействие РНКП и PHKI. [10] |
Критическим событием инициации репликации плазмид этого типа является комплементарное спаривание РНКП с ДНКориджи-на. Естественно, именно эта стадия инициации репликации и регулируется. [11]
Фундаментальная основа передачи информации с помощью нуклеиновых кислот - комплементарное спаривание оснований - выглядит настолько изящной и величественной, что мы на время оказались ослепленными ею и неспособными к дальнейшему совершенствованию наших представлений о структуре ДНК. Но постепенно пелена с глаз спадает, и уже многие лаборатории в мире пытаются добавить свой мазок к будущей более точной картине. [12]

Доказательства того, что включение аминокислоты в полипептидную цепь определяется комплементарным спариванием оснований между кодоном мРНК и антикодоном тРНК, а не самой аминокислотой, были получены в следующем эксперименте. При воздействии на этот комплекс катализатора - никеля Ренея - цистеин превращается в аминокислоту аланин. Когда такой новый комплекс тРНК - аланин ( несущий антикодон тРНК - цистеина) помещали в бесклеточную систему, содержавшую поли ( УГУ) - мРНК, синтезированная поли-пиптидная цепь состояла только из аланина. Этот эксперимент пвдтвердил важную роль механизма образования комплекса мРНК - ко-дон - тРНК - антикодон в трансляции генетического кода. [14]
Эффективность терминации зависит от прочности терминагор-ной шпильки в РНК - Это видно из того, что мутации, приводящие к нарушению комплементарного спаривания какого-либо основания в шпильке, ослабляют терминацию, а мутации, восстанавливающие комплементарность, усиливают терминацию. Усиление прочности РНК-ДНК гибрида в районе олигои ослабляет терминацию. [15]
Страницы: 1 2 3 4