Сплайсинг
Cтраница 1
Сплайсинг рассматривался на примерах превращения одной молекулы РНК - Однако принципиальный интерес представляет обнаружение случаев транс-сплайсинга, когда происходит образование ко-валентных связей между фрагментами РНК, синтезированными на разных генах. Эти гены могут быть локализованы даже в разных: хромосомах. Вероятно, этот процесс облегчается в результате установления временных комплементарных взаимодействий между разными молекулами РНК - Транс-сплайсинг хорошо показан у паразитических одноклеточных эукариот - трипанозом. Мини-экзоны и экзоны тела мРНК могут располагаться в разных: хромосомах. [1]
Сплайсинг рассматривался на примерах превращения одной молекулы РНК - Однако принципиальный интерес представляет обнаружение случаев транс-сплайсинга, когда происходит образование ко-валентных связей между фрагментами РНК, синтезированными на разных генах. Эти гены могут быть локализованы даже в разных хромосомах. Вероятно, этот процесс облегчается в результате установления временных комплементарных взаимодействии между разными молекулами РНК. Транс-сплайсинг хорошо показан у паразитических одноклеточных эукариот - трипанозом. Мини-экзоны и экзоны тела мРНК могут располагаться в разных хромосомах. [2]

Иногда сплайсинг мРНК может проходить по альтернативному варианту. [4]
Такой сплайсинг, называемый альтернативным, имеет большое значение в регуляции транскрипции. [5]
Нарушения сплайсинга могут быть губительны для организма. У человека известна аутосомная рецессивная мутация в гене фени-лаланингидроксилазы. В гомозиготном состоянии мутация приводит к заболеванию - фенилкетонурии, проявляющейся в резкой задержке развития и смерти в раннем детском возрасте при отсутствии специальной диеты. Отсутствие фермента, катализирующего гидроксилирование фенилаланина с образованием тирозина, приводит к накоплению фенилаланина и нарушению образования многих метаболитов, образующихся из ароматических аминокислот. Единственная замена в 5 -районе 12-го интрона, превращающая канонический динуклео-тид GT в AT, приводит к нарушению сплайсинга и процессинга. Образующаяся аберрантная РНК, не содержащая концевого экзо-на, служит матрицей для трансляции преждевременно обрывающегося полипептида, который быстро распадается в клетке. Нарушения сплайсинга описаны также для генов человека, кодирующих глобины. [6]
Нарушения сплайсинга могут быть губительны для организма. У человека известна аутосомная рецессивная мутация в гене фени-лаланннгидроксилазы. В гомозиготном состоянии мутация приводит к заболеванию - фенилкегонурии, проявляющейся в резкой задержке развития и смерти в раннем детском возрасте при отсутствии специальной диеты. Отсутствие фермента, катализирующего гидроксилирование фенилаланина с образованием тирозина, приводит к накоплению фенилаланина и нарушению образования многих метаболитов, образующихся из ароматических аминокислот. Единственная замена в 5 -районе 12-го нитрона, превращающая канонический динуклео-тид GT в AT, приводит к нарушению сплайсинга и процессинга. Образующаяся аберрантная РНК, не содержащая концевого экзо-на, служит матрицей для трансляции преждевременно обрывающегося полнпептида, который быстро распадается в клетке. Нарушения сплайсинга описаны также для генов человека, кодирующих глобины. [7]
Нарушения сплайсинга могут быть губительны для организма. У человека известна аутосомная рецессивная мутация в гене фени-лаланингидроксилазы. [8]
Явление сплайсинга РНК вполне обычно для эукариотических клеток, т.е. клеток, имеющих ядро. Считают, что для прокариотических клеток, не содержащих отчетливо выраженных ядер, сплайсинг отсутствует. [9]
Ферментативная машина сплайсинга, локализованная в ядре, изучена недостаточно, однако ряд важных результатов получен при исследовании превращений предшественников индивидуальных мРНК ( например, р-глобина) в ядерных экстрактах in vitro. Прежде всего происходит разрыв нуклеотидной последовательности на границе экзона 1 и интрона с образованием 5 -конца, представленного фос-форилированным гуанозином - каноническим нуклеотидом левого ( 5 -конца) интрона. Следующая ( или сопряженная с предыдущей) реакция заключается во взаимодействии 5 -конца удаляемой последовательности с 2 -гидроксилом аденозина интрона с образованием; 2 - 5 -фосфодиэфирной связи. Образование такой структуры можно также рассматривать как результат реакции трансэтерификации, в результате которой 2 - ОН-группа аденозина атакует фосфодиэфирную связь, замещая З - ОН-группу экзона 1 Теперь экзон 1 представлен отдельной молекулой РНК, удерживаемой вместе с другой молекулой, содержащей лассд-интрон и экзон 2, в едином комплексе с белками, участвующими в реакции. По-видимому, образование лассо является условием для завязывания ковалентной связи З - ОН-группы линейной молекулы экзона 1 с экзоном 2 в районе сайта сплайсинга; эта реакция также представляет собой трансэтерификацию, когда З - ОН-группа экзона 1 атакует фосфодиэфирную связь и замещает З - ОН-группу интрона; последовательность интрона выщепляется из РНК в форме лассо. В результате число фосфодиэфирных связей в составе субстрата и продукта не изменяется. [10]
Ферментативная машина сплайсинга, локализованная в ядре, изучена недостаточно, однако ряд важных результатов получен при исследовании превращений предшественников индивидуальных мРНК ( например, р-глобина) в ядерных экстрактах in vitro. Прежде всего происходит разрыв нуклеотидной последовательности на границе экзона 1 и интрона с образованием 5 -конца, представленного фос-форилированным гуанозином - каноническим нуклеотидом левого ( 5 -конца) интрона. Следующая ( или сопряженная с предыдущей) реакция заключается во взаимодействии 5 -конца удаляемой последовательности с 2 -гидроксилом аденозина интрона с образованием. Образование такой структуры можно также рассматривать как результат реакции трансэтерификации, в результате которой 2 - ОН-группа аденозина атакует фосфодиэфирную связь, замещая З - ОН-группу экзона 1 Теперь экзон 1 представлен отдельной молекулой РНК, удерживаемой вместе с другой молекулой, содержащей лассд-интрон и экзон 2, в едином комплексе с белками, участвующими в реакции. По-видимому, образование лассо является условием для завязывания ковалентной связи З - ОН-группы линейной молекулы экзона 1 с экзоном 2 в районе сайта сплайсинга; эта реакция также представляет собой трансэтерификацию, когда З - ОН-группа экзона 1 атакует фосфодиэфирную связь и замещает З - ОН-группу интрона; последовательность интрона выщепляется из РНК в форме лассо. В результате число фосфодиэфирных связей в составе субстрата и продукта не изменяется. [11]
Ферментативная машина сплайсинга, локализованная в ядре, изучена недостаточно, однако ряд важных результатов получен при исследовании превращений предшественников индивидуальных мРНК ( например, р-глобина) в ядерных экстрактах in vitro. Прежде всего происходит разрыв нуклеотидной последовательности на границе экзона 1 и интрона с образованием 5 -конца, представленного фосфор ил ированным гуанозином - каноническим нуклеотидом левого ( 5 -конца) интрона. Следующая ( или сопряженная с предыдущей) реакция заключается во взаимодействии 5 -конца удаляемой последовательности с 2 -гидроксилом аденозина интрона с образованием; 2 - 5 -фосфодиэфирной связи. Образование такой структуры можно также рассматривать как результат реакции трансэтерификации, в результате которой 2 - ОН-группа аденозина атакует фосфодиэфирную связь, замещая З - ОН-группу экзона 1 Теперь экзон 1 представлен отдельной молекулой РНК, удерживаемой вместе с другой молекулой, содержащей лассб-интрон и экзон 2, в едином комплексе с белками, участвующими в реакции. По-видимому, образование лассо является условием для завязывания ковалентной связи З - ОН-группы линейной молекулы экзона 1 с экзоном 2 в районе сайта сплайсинга; эта реакция также представляет собой трансэтерификацию, когда З - ОН-группа экзона 1 атакует фосфодиэфирную связь и замещает З - ОН-группу интрона; последовательность интрона выщепляется из РНК в форме лассо. В результате число фосфодиэфирных связей в составе субстрата и продукта не изменяется. [12]
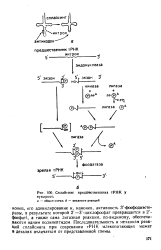 |
Сплайсинг предшественника тРНК у. [13] |
Последовательность и механизм реакций сплайсинга при созревании тРНК млекопитающих может в деталях отличаться от представленной схемы. [14]
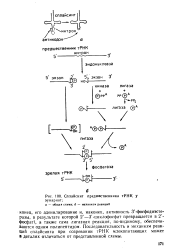 |
Сплайсинг предшественника тРНК У. [15] |
Страницы: 1 2 3