Спорогония
Cтраница 2
Значение каждой из перечисленных процедур различно для определения заражения, но они имеют свои критерии и свой принципиальный смысл. С помощью вскрытия и просмотра нативного материала под покровным стеклом устанавливаем локализацию патологических процессов в тканях, изменения в органах и стадии, которые появляются в спорогонии паразита, главным образом количество спор в панспоробласте. На сухих мазках обнаруживаем преимущественно главные типы стадий и их ядерные соотношения. [16]
Шизогрегарины могут быть заметно патогенны для насекомых-хозяев. Представители этой группы размножаются как спорогонией, так и шизогонией. Спорогония происходит так же, как и у эугрегарин. Шизогония может происходить вне или внутри клетки хозяина, осуществляется путем двойного деления, множественного деления и почкования. Из 25 - 30 видов шизогрегарин, выделенных из насекомых, большинство было найдено у жуков, клопов и двукрылых. Если из слияния двух гаметоцитов образуется одна спора, шизогрегарина относится к семейству Ophryocystidae; если же образуются две споры или больше, она принадлежит к Schizocystidae. [17]
Кокцидии этого рода чаще всего встречаются в губоногих и двупарноногих, но также и в дождевых червях; лишь небольшое число видов найдено в насекомых. Legerella hydroporini Vincent развивается в мальпигиевых сосудах водолюба Hydroporus palu-stris. Схизогония и спорогония у этого вида проходят в клетках эпителия сосудов, и лишь созревшие споры, освобождаясь, поступают в канал сосуда, затем в полость кишечника и с экскрементами выделяются наружу. Макрогаметы яйцевидные, вакуолизи-рованные, с пузыревидным ядром и крупной кариосомой. Микрогаметы яйцевидные, в 10 раз меньше макрогамет, к которым они прикрепляются. Зигота заключена в бесформенно изогнутой оболочке, в ней происходит многократное деление ядра. В конечном итоге дифференцируется большое количество булавовидных спо-розоитов, концы которых лежат в остаточном теле. Циста одновременно является и спорой. [18]
У микроспоридий подобные проблемы встречаются гораздо чаще. Это можно хорошо продемонстрировать на видах рода Thelohania; его определяют таким образом, что в спорогонии у представителей рода образуется восьмиядерный панспоробласт. В типичном случае в спорогонии постепенно образуются 2 -, 4 - и 8-ядерные стадии. Вокруг ядер 8-ядерной стадии обособляются пальцевидные, овальные споробласты, которые превращаются в овальные споры, удерживаемые вместе мембраной - стенкой панспоробласта. Восьмерка спор отчетливо различается и спустя долгое время после гибели хозяина. Примером может служить Thelohania pinguis у комаров хирономусов. В воде эти плазматические остатки набухают, ц споры расходятся. [19]
Листья одуванчика, например, в сравнении с листьями клеверов и подорожника значительно задерживали спорообразование микроспоридий. При содержании гусениц на питательных средах, в состав которых входили антисептики и антибиотики, микроспоридии позже приступали к спорогонии, а панспоробласты содержали мало спор. [20]
В период спорогонии важное значение имеет температура внешней среды. От этого в значительной степени зависит продолжительность спорогонии. Допустимым нижним температурным порогом является температура 16 С, верхним - 30 С. В других условиях спорогония не происходит. [21]
Большинство видов гемоспоридий не являются настоящими паразитами, весь жизненный цикл которых связан с насекомыми и для них патогенных. Для определения видовой принадлежности разных видов гемоспоридий в позвоночных животных имеются хотя бы некоторые морфологические данные, однако о стадиях этих простейших, развивающихся в насекомых, такие данные почти полностью отсутствуют. Приводимые ниже литературные данные ( часто без указания первоисточников) могут служить лишь введением к широким исследованиям, которые еще предстоят про-тозоологам и энтомопатологам. Не вдаваясь в подробности биологического цикла развития гемоспоридий в позвоночных животных, следует указать, что для большинства видов имеют место три типичных цикла: схизогония, гамогония и спорогония. В позвоночных животных гемоспоридий проходят преимущественно цикл схизогонии как в кровяных тельцах, так и в ретикулярных тканях различных органов. Гамогония гемоспоридий в позвоночных животных протекает лишь до образования гаметоцитов, циркулирующих в периферийных кровеносных сосудах и всасываемых с кровью насекомыми-переносчиками, в органах которых гамогония заканчивается и начинается спорогония. [22]
У семенных растений микроспоре гомологично пыльцевое зерно. У покрытосеменных ( цветковых) микроспорангию гомологично гнездо пыльника. Спорогонии предшествует половой процесс - кариогамия. У разных родов из одного споронга образуется разл. [23]
При спорогонии из споронта возникает споробласт ( у видов с одной спорой) или из плазмодия развивается панспоробласт со многими спорами. Плазмодии сохраняют оболочку, в которую они заключены и в тот период, когда их содержимое дифференцировалось на множество одноядерных споробластов. Споробласты шаровидные, позднее становятся удлиненными, и на их поверхности образуется прочная, стойкая оболочка споры. У многих видов микроспоридий оболочка панспоробластов подвергается автолизу, и споробласты или споры освобождаются и располагаются в беспорядке. Такие виды здесь исключены из рода Nosema и отнесены к родам, которым свойственны формы, недоразвитые в стадии спорогонии, еще способные удерживаться в группах. [24]
Большинство видов гемоспоридий не являются настоящими паразитами, весь жизненный цикл которых связан с насекомыми и для них патогенных. Для определения видовой принадлежности разных видов гемоспоридий в позвоночных животных имеются хотя бы некоторые морфологические данные, однако о стадиях этих простейших, развивающихся в насекомых, такие данные почти полностью отсутствуют. Приводимые ниже литературные данные ( часто без указания первоисточников) могут служить лишь введением к широким исследованиям, которые еще предстоят про-тозоологам и энтомопатологам. Не вдаваясь в подробности биологического цикла развития гемоспоридий в позвоночных животных, следует указать, что для большинства видов имеют место три типичных цикла: схизогония, гамогония и спорогония. В позвоночных животных гемоспоридий проходят преимущественно цикл схизогонии как в кровяных тельцах, так и в ретикулярных тканях различных органов. Гамогония гемоспоридий в позвоночных животных протекает лишь до образования гаметоцитов, циркулирующих в периферийных кровеносных сосудах и всасываемых с кровью насекомыми-переносчиками, в органах которых гамогония заканчивается и начинается спорогония. [25]
Виды двух первых отрядов, встречающиеся в насекомых, имеют сходный цикл развития, который можно разделить на период схизогонии и период гамогонии и спорогонии. Схизогония в обоих отрядах происходит в тканях хозяина, в цитоплазме пораженных клеток. Из зародыша, проникшего в тело хозяина, созревает схи-зонт, вырастающий в шаровидную или удлиненную стадию с несколькими ядрами, вновь распадающуюся на одноядерные мерон-ты. Процесс схизогонии у большинства видов повторяется до тех пор, пока паразит не размножится в такой степени, что приобретет господствующее положение в теле хозяина. В определенный момент структура сформировавшихся стадий изменяется, и возникают обычные формы с пузыревидным ядром, в котором хроматин расположен в хромосомах. Возникают раздельные изо-гаметы, что мы предполагаем у гаплоспоридий, или образуются двуядерные формы, известные как диплокарионы, где гаметы представлены лишь дифференцированными ядрами одной и той же клетки, которые сливаются и превращаются в зиготу, а из нее в дальнейшем развиваются споронты. В процессе спорогонии после одного или нескольких делений образуются споробла-сты, которые, созревая, дают споры с прочной оболочкой. Споры монолитны, у некоторых видов ( Haplosporidia - из организма червей) имеют поры. Внутри споры обычно имеется зародыш, форма которого соответствует самой споре. [26]
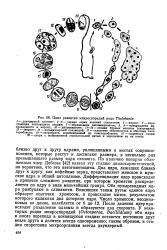 |
Цикл развития микроспорйдий рода Thelohania. [27] |
По наличию попарно сближенных ядер Дебезье [42] назвал эту стадию диплокариотической, но вероятнее, что это автогаметоциты. Два ядра, лежащих близко друг к другу, как кофейные зерна, представляют женское и мужское ядра в общей цитоплазме. Дифференциация ядер происходит в процессе сложного развития, при котором хроматин распределяется по ядру в виде мельчайших гранул. Оба пузыревидных ядра разбухают и сливаются. Возникшее таким путем единое ядро образует ленточки хромосом, и происходит митоз, результатом которого являются два дочерних ядра. После деления ядер происходит разделение плазмы и образуются два споронта. У некоторых родов микроспоридий ( Ociosporea, Bacillidium) оба ядра и после перехода в покоящуюся стадию остаются постоянно друг возле друга и делятся всегда одновременно; таким образом, в стадии спорогонии микроорганизм всегда двуядерный. [28]
Страницы: 1 2