Стадия - инициация
Cтраница 2
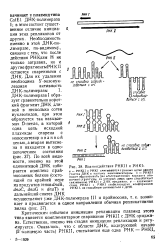
Критическим событием инициации репликации плазмид этого типа является комплементарное спаривание РНКИ с ДНК ориджи-на. Естественно, именно эта стадия инициации репликации и регулируется. [17]
 |
Взаимодействие РНКП и PHKI. [18] |
Критическим событием инициации репликации плазмид этого типа является комплементарное спаривание РНКП с ДНКориджи-на. Естественно, именно эта стадия инициации репликации и регулируется. [19]
Установлено, что этот фермент содержит специфический центр, в котором связывается рифамицин. Рифамицин оказывает свое действие на стадии инициации, препятствуя образованию первой межнуклеотидной связи. Другие вещества ингибируют элонгацию, влияя на РНК-полимеразу. [20]
 |
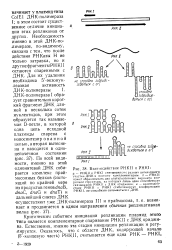
Нуклеотидная последовательность концевых участков ( - J - и ( - цепей РНК фага Qfi. [21] |
Однако при наличии затравки фермент может копировать любую РНК: таким образом, специфичность проявляется на стадии инициации. Любопытно, что избирательность фермента по отношению к матрице в значительной степени определяется входящими в его состав клеточными белками: белок S1 и хозяйский фактор требуются при использовании в качестве матрицы ( -) нити РНК фага Qp, но эти белки ненужны, когда в роли матрицы выступает, например, ( -) нить этой РНК. [22]
 |
Нуклеотидная последовательность концевых участков ( и ( - цепей РНК фага Q. [23] |
Однако при наличии затравки фермент может копировать любую РНК: таким образом, специфичность проявляется на стадии инициации. Любопытно, что избирательность фермента по отношению К матрице в значительной степени определяется входящими в его состав клеточными белками: белок S1 и хозяйский фактор требуются при использовании в качестве матрицы ( -) нити РНК фага Qp, но эти белки ненужны, когда в рати матрицы выступает, например, ( -) нить этой РНК. [24]
 |
Нуклеотидная последовательность концевых участков ( - J - и ( - цепей РНК фага Qp. [25] |
Однако при наличии затравки фермент может копировать любую РНК: таким образом, специфичность проявляется на стадии инициации. Любопытно, что избирательность фермента по отношению к матрице в значитетьной степени определяется входящим в его состав клеточными белками: белок S1 и хозяйский фактор требуются при использовании в качестве матрицы ( -) нити РНК фага Qp, но эти белки ненужны, когда в роли матрицы выступает, например, ( -) нить этой РНК. [26]
Применение двух - и многослойных сталей и сплавов, обладающих взаимодополняющими физико-механическими свойствами, позволяет значительно снизить металлоемкость элементов конструкций. Проблема проектирования, создания и эксплуатации биметаллических конструкций повышенного ресурса, в частности высоконагруженного оборудования АЭС, делает весьма актуальными экспериментальные исследования, направленные на разработку методов оценки несущей способности таких конструкций не только по интегральным характеристикам прочности, но и с учетом наличия трещиноподоб-ных дефектов на стадиях инициации разрушения, а также распространения и остановки трещин. Развитие методов определения критериев сопротивления разрушению и их анализ необходимы для оптимизации свойств биметалла путем правильного выбора сочетания разнородных составляющих соединения, назначения технологического способа его изготовления и определения рационального соотношения толщин основного металла и плакирующего слоя. Кроме того, это необходимо при проведении расчетов на прочность и оценке ресурса биметаллических элементов конструкций, определении допускаемых размеров дефектов, выборе методов и средств дефектоскопии. [27]
Первая стадия ( инициация) начинается с присоединения матричной РНК ( мРНК) к малой рибосомной субчастице, не связанной с большой субчастицей. В стадии инициации участвуют спец. Генетический код), инициаторная транспортная РНК ( тРНК) и специфич. [28]
Следующая стадия, инициация, требует наличия субстратов РНК-полимеразы, нуклеозидтрифосфатов и заключается в образовании первых нескольких звеньев цепи РНК. Первый нуклеотид входит в состав цепи, сохраняя свою трифосфатную группу, а последующие присоединяются к З - ОН-группе предыдущего с освобождением пирофосфата. На стадии инициации РНК-продукт связан с матрицей и РНК-полимеразой непрочно и с высокой вероятностью может освобождаться из комплекса. В этом случае РНК-полимераза, не покидая промотора, снова инициирует РНК. Когда РНК-продукт достигает критической длины ( от 3 до 9 нуклео-тидов на разных промоторах), абортивная инициация полностью прекращается, транскрибирующий комплекс стабилизируется и уже не распадается до тех пор, пока синтез молекулы РНК не будет доведен до конца. Примерно в этот же момент, который считается концом инициации и началом элонгации, от РНК-полимеразы отделяется т-субъединица. [29]
ДНК-зависимый синтез РНК подавляется рядом антибиотиков, таких как актиномицин, которые блокируют матрицу ДНК. Антибиотики группы рифамицина, напротив, действуют на РНК-полимеразу. Рифамицин действует на стадии инициации, препятствуя образованию первой межнуклеотидной связи. Другие вещества, влияющие на полимеразу, ингибируют элонгацию. [30]
Страницы: 1 2 3