Тилакоид
Cтраница 2
Во внутренней мембране тилакоидов размещены партикулы размером около 17 нм, выступающие над поверхностью мембраны в полость камеры тилакоида. [16]
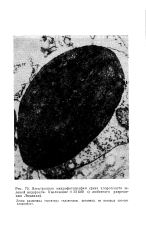 |
Электронная микрофотография среза хлоропласта зеленой водоросли. Увеличение X 33 600 ( с любезного разрешения Людвика. [17] |
Легко различима структура тилакоидов, органелл, из которых состоит хлоропласт. [18]
 |
Схематическое изображение ультраструктуры мембран тилакоидов, собранных в граны, и мембран одиночных тилакоидов. [19] |
Структура мембраны как тилакоидов гран, так и тила-коидов стромы ( одиночных тилакоидов) была подробно изучена с помощью электронной микроскопии и иммунологических методов. Много информации получено также и о химическом составе этих мембран. [20]
Образование хроматофоров ( тилакоидов индивидуальных и их скоплений) происходит только при росте бактерий в присутствии света. [21]
Большую часть цитоплазмы занимают тилакоиды, располагающиеся обычно концентрическими кругами по периферии клетки. Тилакоиды, как и у цианобактерий, лежат в цитоплазме свободно, т.е. не отделены от нее замкнутой мембраной, и имеют тенденцию сближаться, образуя пары или стопки, состоящие из трех и более тилакоидов. Внутренние и наружные поверхности тилакоидных мембран гладкие. Фикобилисомы и фикобилипроте-ины не обнаружены. [22]
Изменение концентрации протонов внутри тилакоидов регулирует скорость переноса электронов между ФС1 и ФСП. [23]
Регуляция синтеза пигментов и тилакоидов. Синтез фотопигментов зависит от условий роста организмов, прежде всего от освещенности и ( у факультативных анаэробов) от присутствия кислорода. [24]
Фотохимический центр ФС1 пронизывает всю мембрану тилакоида насквозь так, что реакционный центр Рт находится на внутренней стороне мембраны. Акцептор электронов А, по-видимому, содержит Fe - S-белки и целую систему белков, связанных с Jep-редоксином. Он расположен на внешней стороне мембраны. [25]
Таким образом, в слоевых системах тилакоидов имеются сложные пигментно-липидно-белковые комплексы с различными рассмотренными выше простетическими группами; только оптимальная пространственная организация этих комплексов делает возможным столь быстрый и эффективный транспорт электронов по цепи переносчиков, который наблюдается в фотосинтезе. Однако та же пространственная организация, вероятно, предопределяет и участие тех или иных компонент в нескольких редокс-системах, и возникновение новых, многокомпонентных редокс-систем, которое стимулируется условиями внешней среды живого организма, в частности действием мутагенов, ингибиторов и других агентов. Например, пластохинон А - первый акцептор электрона от Хл реакционных центров фотосистемы II - является еще и кофактором циклического переноса электрона с участием только системы I. Имеются данные о том, что цитохром f - важное звено в цепи транспорта электрона от фотосистемы II к фотосистеме I - принимает участие и в циклическом транспорте электрона. [26]
От этих кармашков отшнуровываются плоские пузырьки - тилакоиды, постепенно увеличивающиеся в размерах ( фиг. Образовавшиеся таким способом в темноте из инициальных частиц органоиды с диаметром 0 5 - 0 8 мк, видимые в оптический микроскоп, получили название пропластид. [27]
Сама фотосинтетическая мембрана, из которой образованы тилакоиды, состоит из бимолекулярного липидного слоя, частично или полностью пронизывающего белковые макромолекулы. Липи-ды находятся в аморфном, а не в кристаллическом состоянии, как считалось ранее. [28]
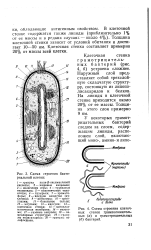 |
Схема строения бактериальной клетки.| Схема строения клеточных стенок грамположитель-ных ( а и грамотрицательных ( б бактерий. [29] |
S - включения серы; 4 - трубчатые тилакоиды; 5 - пластинчатые тила-коиды; 6 - пузырьки; 7 - хромато-форы; 8 - ядерное вещество ( ну-клеоид); 9 - рибосомы; 10 - цитоплазма; / / - базальное тельце; 12 - жгутики; 13 - капсула; 14 - клеточная стенка; 15 - цитоплаз-матическая мембрана; 16 - мезосо-ма; 17 - газовые вакуоли; 18 - ла-меллярные структуры; 19 - гранулы полисахарида; 20 - гранулы полифосфата. [30]
Страницы: 1 2 3 4 5