Тироглобулин
Cтраница 1
Тироглобулин - основной компонент коллоида фолликулов - является гликопротеидом с молекулярной массой 660000, состоящим из четырех идентичных субъединиц, содержит 0 5 - 1 % йода и 8 - 10 % углеводов. Он представляет собой запасную форму тиреоидных гормонов и содержит обычно 90 % от общего количества йода, присутствующего в щитовидной железе. Количество и соотношение различных форм йода в щитовидной железе зависят от множества факторов - скорости поступления в нее йода, присутствия струмогениых веществ - гойтерогенов, которые могут нарушить механизм улавливания йода, от некоторых патологических состояний, включая влияние генетических факторов. [1]
Тироглобулин представляет собой сложный гликопротеин, содержащийся в фолликулах щитовидной железы и являющийся средством хранения ( в связанной форме) двух гормонов щитовидной железы: тироксина ( 51) и трииодтиронина. На основании имеющихся данных [64] можно предположить, что биосинтез тироксина протекает по механизму окислительного сочетания фенолов из двух определенным образом расположенных остатков дииод - L-тирозина в белковой молекуле тироглобулина. Однако возможны и иные механизмы. Истинная роль тироглобулина в окислительном связывании недостаточно выяснена. Известно, что иодирование остатков L-тирозина других полипептидов легко происходит ш vitro, однако образование тироксина ( 51) in vivo наблюдается только при иодировании тироглобулина. [2]
 |
Тиреоидные гормоны ( А и Б и мо-ноиодтирозин ( В-первый иодированный предшественник тиреоидных гормонов. [3] |
Почти весь новообразованный тироксин и трииодтиронин сохраняют связь с тироглобулином в тиреоидных пузырьках до тех пор, пока они не поступят в кровь, высвободившись под действием протеолитических ферментов. Полагают, что тироксин является предшественником, или прогор-моном, трииодтиронина, стимулирующего обмен веществ. [4]
Рош [6] описал способ выделения З - йодтирозина - J131 и 3, 5-ди-подтирозина - J 31 из гидролизатов тироглобулина методом бумажной хроматографии ( ватман № 1); хроматограмму проявляют смесью, содержащей 68 % бутанола, 5 % уксусной кислоты и 27 % воды. [5]
Расположите события, происходящие при синтезе и секреции тироидных гормонов, в необходимом порядке: 1) иодирование остатков тирозина в тироглобулине; 2) синтез тиролибе - рина в гипоталамусе; 3) объединение двух иодированных остатков молекул тирозина с образованием три - и тетраиодпро-изводных; 4) поступление иодтиронинов в клетки-мишени; 5) синтез и секреция тиротропина; 6) связывание иодтиронинов с транспортными белками. [6]
I), метилтиоурацил ( II), угнетают активность ферментных систем, участвующих в окислении иодидов в иод, что приводит к торможению иодирования тироглобулина и, следовательно, к задержке образования моно - и дииодтирозина, а также превращения последних в тироксин и трииодтиро-нин. [7]
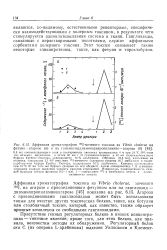 |
Аффинная хроматография 1251-меченого токсина из Vibrio cholerae на фетуин-агарозе ( а и на ганглиозиддиаминодипропиламино-агарозе ( б. [8] |
Ганглиозиды, связанные с нерастворимыми носителями, служат аффинными сорбентами холерного токсина. Этот токсин связывает также некоторые гликолротеины, такие, как фетуин и тироглобулин. [9]
Тироглобулин представляет собой сложный гликопротеин, содержащийся в фолликулах щитовидной железы и являющийся средством хранения ( в связанной форме) двух гормонов щитовидной железы: тироксина ( 51) и трииодтиронина. На основании имеющихся данных [64] можно предположить, что биосинтез тироксина протекает по механизму окислительного сочетания фенолов из двух определенным образом расположенных остатков дииод - L-тирозина в белковой молекуле тироглобулина. Однако возможны и иные механизмы. Истинная роль тироглобулина в окислительном связывании недостаточно выяснена. Известно, что иодирование остатков L-тирозина других полипептидов легко происходит ш vitro, однако образование тироксина ( 51) in vivo наблюдается только при иодировании тироглобулина. [10]
Синтез и секреция Т3 и Т4 контролируются тиролиберином и тиротропином. Тиротропин оказывает следующие эффекты: стимулирует активный транспорт 1 - против 500-кратного градиента в полость фолликула за счет цАМФ - зависимого фосфорилирования белков клеточных мембран; усиливает транскрипцию и трансляцию тироглобулина; стимулирует рост эпителиальных клеток, формирующих фолликулы, а в фолликулярном коллоиде - иодирование тирозилов; по аденилатциклазному механизму стимулирует синтез Т3, Т4 ( аналогично действуют адреналин и PGE2); стимулирует секрецию иодированного тироглобулина путем пиноцитоза и отщепления Т3 и Т4 при слиянии пиноцитозных пузырьков с мембранами лизосом ( протеолитическим путем), а также поступление Т3 и Т4 в кровь и лимфу. [11]
Синтез и секреция Т3 и Т4 контролируются тиролиберином и тиротропином. Тиротропин оказывает следующие эффекты: стимулирует активный транспорт 1 - против 500-кратного градиента в полость фолликула за счет цАМФ - зависимого фосфорилирования белков клеточных мембран; усиливает транскрипцию и трансляцию тироглобулина; стимулирует рост эпителиальных клеток, формирующих фолликулы, а в фолликулярном коллоиде - иодирование тирозилов; по аденилатциклазному механизму стимулирует синтез Т3, Т4 ( аналогично действуют адреналин и PGE2); стимулирует секрецию иодированного тироглобулина путем пиноцитоза и отщепления Т3 и Т4 при слиянии пиноцитозных пузырьков с мембранами лизосом ( протеолитическим путем), а также поступление Т3 и Т4 в кровь и лимфу. [12]
Йодированный тиро-глобулин и тиреоидные гормоны подвергаются реабсорб-ции эпителиальными клетками фолликулов, где происходят их протеолиз и частичная деградация. Ббльшая часть тиреоидных гормонов при этом сохраняется и секретиру-ется в кровеносное русло [ Wullman S. При гидролизе тироглобулина образуется также йодированное производное L-гистидина. Щитовидная железа способна к реутилизации йода. Она содержит микросо-энальную дегалогеназу, зависящую от NADP, катализирующую дейодирование MIT и DIT, не действует на йодтиронины. [13]
Выделяемый базофильными клетками передней доли гипофиза тиротропный гормон ( ТТГ) стимулирует функцию щитовидной железы. Механизмы этой стимуляции многообразны. Активируя протеазы, ТТГ повышает распад тироглобулина в щитовидной железе, что приводит к усиленному выделению тироксина и трийодтиронина в кровь. ТТГ способствует накоплению йода в щитовидной железе; кроме того, он повышает активность ее секреторных клеток и увеличивает их число. [14]
Гормоны щитовидной железы синтезируются в составе тирогло-булина. Тироидная йодная помпа связана с Na, К - АТФазой. Затем происходит электрофильная атака тирозиновых радикалов белка тироглобулина и образуются моноиод - и дииодтирозилы ( MIT-TG, DIT-TG), которые сшиваются в трииодтиронилы и тироксилы. Иодированные молекулы тироглобулина посредством пиноцитоза поступают в клетки фолликулов. [15]
Страницы: 1 2