Тироглобулин
Cтраница 2
Гальми и Штюльке57 сообщили, что в то время как подкожное введение 100 мг или более перхлората натрия мешает активному потреблению J131 щитовидной железой крысы, получавшей пропилтиоурацил при введении перхлората натрия после приема J131, далеко не весь захваченный J131 выводится из железы. Чем больше интервал между введением J131 и перхлората натрия, тем меньшее количество J131 может быть выведено из железы. Не способный выделяться J131, по-видимому, не может быть связан в виде тироглобулина. [16]
Регуляция: тиролиберин гипоталамуса стимулирует выделение тиротропина. Соматотропин ингибирует выделение тиротропина. Циркулирующие в крови Т3 и Т4 подавляют секрецию тиролиберина и тиротропина; Т4 подавляет синтез тироглобулина. Иодтиронины транспортируются в комплексе с TBG и ТВРА. [17]
 |
Содержание углеводов в некоторых гликопротеинах. [18] |
По-видимому, в природе существует гораздо большее число белков, ковалентно связанных с углеводами, чем чистых белков. Гликопротеины широко распространены в тканях высших животных, растениях и микроорганизмах и выполняют различные функции. Так, структурными гликопротеинами являются коллаген, гликопротеины клеточной стенки бактерий, экстензии из клеточной стенки растений; резервную функцию выполняют казеин, овальбу-мин, гликопротеины эндосперма, аллергены цветочной пыльцы; ферментами являются рибонуклеаза В свиньи, дезоксирибонук-леаза свиньи, а-амилаза свиньи, ацетилхолинэстераза, фицин, бро-мелаин, такаамилаза ( а-амилаза грибов), дрожжевая инвертаза ( p - D-фруктофуранозидаза); транспортную функцию выполняют церулоплазмин, гаптоглобулин, трансферрин; гормонами служат тироглобулин, хорионический гонадотропный гормон ( хориониче-ский гонадотропин) человека, фолликулостимулирующий гормон, эритропоэтин; защитную функцию выполняют фибриноген, интерферон, иммуноглобулины, муцины; в плазме и других жидкостях организма содержатся а -, 5 - и - гликопротеины; рицин, фито-токсины грибов являются токсинами. Гликопротеины из разных источников различаются содержанием углеводов ( табл. 26.3.4 J, а также размером и формой углеводных цепей. Роль углеводных остатков в гликопротеинах ясна не до конца. [19]
Гормоны щитовидной железы синтезируются в составе тирогло-булина. Тироидная йодная помпа связана с Na, К - АТФазой. Затем происходит электрофильная атака тирозиновых радикалов белка тироглобулина и образуются моноиод - и дииодтирозилы ( MIT-TG, DIT-TG), которые сшиваются в трииодтиронилы и тироксилы. Иодированные молекулы тироглобулина посредством пиноцитоза поступают в клетки фолликулов. [20]
Тироглобулин представляет собой сложный гликопротеин, содержащийся в фолликулах щитовидной железы и являющийся средством хранения ( в связанной форме) двух гормонов щитовидной железы: тироксина ( 51) и трииодтиронина. На основании имеющихся данных [64] можно предположить, что биосинтез тироксина протекает по механизму окислительного сочетания фенолов из двух определенным образом расположенных остатков дииод - L-тирозина в белковой молекуле тироглобулина. Однако возможны и иные механизмы. Истинная роль тироглобулина в окислительном связывании недостаточно выяснена. Известно, что иодирование остатков L-тирозина других полипептидов легко происходит ш vitro, однако образование тироксина ( 51) in vivo наблюдается только при иодировании тироглобулина. [21]
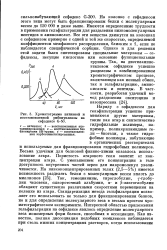 |
Хроматограмма нативной и. [22] |
Пористость агарового геля зависит от концентрации агара. С уменьшением его концентрации в геле доступность внутренних частей зерен для макромолекул увеличивается. Так, гемоцианины, тироглобулин, у-глобу-лин человека, сывороточный альбумин, а - и р-лактоглобулин обладают существенно различными скоростями перемещения на колонке из агара. Стандартизация метода гельфильтрации позволяет его использовать не только для фракционирования белков, но и для определения молекулярного веса в весьма простом хроматографическом эксперименте. [23]
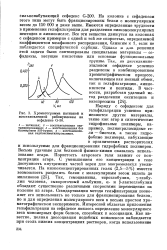 |
Хроматограмма нативной и. [24] |
Пористость агарового геля зависит от концентрации агара. С уменьшением его концентрации в геле доступность внутренних частей зерен для макромолекул увеличивается. Так, гемоцианины, тироглобулин, у-глобу-лин человека, сывороточный альбумин, а - и [ 3-лактоглобулин обладают существенно различными скоростями перемещения на колонке из агара. Стандартизация метода гельфильтрации позволяет его использовать не только для фракционирования белков, но и для определения молекулярного веса в весьма простом хроматографическом эксперименте. [25]
В ходе дальнейшего окисления пероксидазой происходит сопряжение DIT и MIT с образованием различных йодтирони-нов [ Taurog A. Процесс сопряжения, вероятно, происходит следующим образом: из йодированного тирозина образуется радикал с дефицитом электронов, который подвергается - элиминированию с образованием дегидроалани-на и ароматического радикала. Последний реагирует с остатком йодтирозина, связанным с другим участком фермента, образуя йодтиронин ( [ Gavaret J. На 5000 аминокислотных остатков тироглобулина приходится всего 2 - 5 остатков тироксина, что указывает на специфический характер йодирования в участках молекулы белка с достаточно близко расположенными остатками тирозина. [26]
Чтобы определить современное отношение к этому перечню, необходимо, кроме того, различать две группы аминокислот, а именно: аминокислоты, обычно встречающиеся в белках, аминокислоты, встречающиеся только случайно ( иногда) либо в свободном состоянии в организмах высших животных, либо в бактериальных белках. В первом случае метод хроматографии на бумаге позволил определить полный состав большого числа белковых гадролизатов, причем присутствие в таких гидролизатах еще неизвестной до сих пор аминокислоты становится все менее вероятным. Эти авторы не приняли во внимание цистеина, который действительно содержится в целом ряде белков, но включили в список дииодтирозин и тироксин, идентифицированные до сих пор только в тироглобулине. Необходимо, следовательно, уменьшить число аминокислот, первоначально приведенное Викери, до двадцати одного. Поэтому мы должны ограничить перечень двадцатью обычно встречающимися аминокислотами, причем эта цифра никогда, вероятно, не будет значительно изменена. В число 20 аминокислот не включены аспарагин и глутамин, так как они не встречаются в химических белковых гидролизатах. Этот специальный случай будет рассмотрен на стр. [27]
Подобно адреналину, эти два гормона, сокращенно обозначаемые как Т4 и ТЗ соответственно, являются производными тирозина. Тиреоидные гормоны вырабатываются в ответ на сигналы, поступающие в гипоталамус, который секретирует ти-ролиберин, а также ингибитор высвобождения тиротропина. Тиролиберин стимулирует выделение тиротропина из передней доли гипофиза и его поступление в кровь. Тиротропин связывается со специфическими рецепторами на клетках щитовидной железы и стимулирует выработку ими тиреоидных гормонов. Тироксин и трииодтиронин синтезируются в последовательности ферментативных реакций, начинающейся с иодирования остатков L-тирозина в тироглобулине ( гликопротеин с мол. Необходимый для реакции иод, который является незаменимым микроэлементом ( гл. Иод интенсивно связывается и накапливается тирогло-булином, называемым также коллоидным белком, и затем используется для иодирования остатков тирозина в тироглобулине. [28]
Тироглобулин представляет собой сложный гликопротеин, содержащийся в фолликулах щитовидной железы и являющийся средством хранения ( в связанной форме) двух гормонов щитовидной железы: тироксина ( 51) и трииодтиронина. На основании имеющихся данных [64] можно предположить, что биосинтез тироксина протекает по механизму окислительного сочетания фенолов из двух определенным образом расположенных остатков дииод - L-тирозина в белковой молекуле тироглобулина. Однако возможны и иные механизмы. Истинная роль тироглобулина в окислительном связывании недостаточно выяснена. Известно, что иодирование остатков L-тирозина других полипептидов легко происходит ш vitro, однако образование тироксина ( 51) in vivo наблюдается только при иодировании тироглобулина. [29]
Подобно адреналину, эти два гормона, сокращенно обозначаемые как Т4 и ТЗ соответственно, являются производными тирозина. Тиреоидные гормоны вырабатываются в ответ на сигналы, поступающие в гипоталамус, который секретирует ти-ролиберин, а также ингибитор высвобождения тиротропина. Тиролиберин стимулирует выделение тиротропина из передней доли гипофиза и его поступление в кровь. Тиротропин связывается со специфическими рецепторами на клетках щитовидной железы и стимулирует выработку ими тиреоидных гормонов. Тироксин и трииодтиронин синтезируются в последовательности ферментативных реакций, начинающейся с иодирования остатков L-тирозина в тироглобулине ( гликопротеин с мол. Необходимый для реакции иод, который является незаменимым микроэлементом ( гл. Иод интенсивно связывается и накапливается тирогло-булином, называемым также коллоидным белком, и затем используется для иодирования остатков тирозина в тироглобулине. [30]
Страницы: 1 2