Холерный токсин
Cтраница 1
Холерный токсин повышает уровень сАМР в эпителиальных клетках кишечника. Лечение холеры состоит в возмещении потерь жидкости и электролитов в организме. [1]
Холерный токсин, экзотоксин из Vibrio cholerae - белок, ответственный за желудочно-кишечные проявления клинической холеры. Токсин сильно и специфически связывается некоторыми ганглиозидами мозга. [2]
Только с одним типом ганглйозидов вступает в реакцию холерный токсин. Углеводная часть его состоит из цепочки, образованной молекулами трех углеводов - лактозы, галактозамина и галактозы, причем сиаловый радикал присоединен к комлевому, то есть к лактозному компоненту этого сахарного ствола. [3]
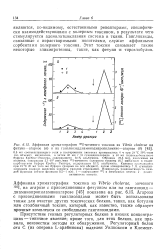 |
Аффинная хроматография 1251-меченого токсина из Vibrio cholerae на фетуин-агарозе ( а и на ганглиозиддиаминодипропиламино-агарозе ( б. [4] |
Ганглиозиды, связанные с нерастворимыми носителями, служат аффинными сорбентами холерного токсина. Этот токсин связывает также некоторые гликолротеины, такие, как фетуин и тироглобулин. [5]
Вибрион холеры вырабатывает гемолитический яд. Холерный токсин обычно представляет собою эндотоксин и освобождается из клетки лишь после ее смерти. [6]
Именно такой подход использовали при создании противохолерной вакцины. Синтетический олигонуклеотид, кодирующий эпитоп субъединицы В холерного токсина, встроили в гипервариабельный участок гена флагеллина Salmonella и полученную конструкцию ввели в дефектный по флагеллину штамм Salmonella. При этом было известно, что эпитоп, включающий 50 - 64 аминокислотные остатки субъединицы В холерного токсина, индуцирует выработку антител к интактному холерному токсину. Химерный флагеллин нормально функционировал, а эпитоп холерного токсина размещался на поверхности жгутиков. Иммунизация мышей с помощью интраперитонеальной инъекции примерно 5 - 106 живых или убитых формалином бактерий с модифицированным флагеллином индуцировала выработку большого количества антител как к пептиду ( 50 - 64 аминокислотным остаткам), так и к молекуле интактного холерного токсина. [7]
При выработке иммунного ответа клеточные рецепторы реагируют на углеводные детерминанты макромолекулы антигена. Обратным примером может служить взаимодействие клеток с макромолекулами холерного токсина. Последний представляет собой белок, в состав которого входят две высокомолекулярные пептидные субъединицы. Одна из них ответственна за первичное взаимодействие с клетками организма-хозяина, а другая - за токсический эффект. Было установлено, что рецептором на поверхности клеток, осуществляющим узнавание молекулы токсина и связывание с ним, является гликолипид - ган-глиозид GMI, в молекуле которого к липидной части присоединен олигосахаридный фрагмент, содержащий остаток сиаловой кислоты. После присоединения токсина к ган-глиозиду от первого отщепляется токсическая субъединица, под дейстием чего происходит ряд изменений в активности ферментов клетки, в первую очередь активация адени-лат-циклазы. [8]
В настоящее время формируется направление, когда в конструкцию вакцин, состоящих из молекул протективных антигенов, ковалентно вшивается участок, обеспечивающий их адгезию на клетках слизистых оболочек. Ими могут служить фрагменты фимбрий энтеробактерий, адгезины вирусов или некоторые субъединииы холерного токсина. [9]
Именно такой подход использовали при создании противохолерной вакцины. Синтетический олигонуклеотид, кодирующий эпитоп субъединицы В холерного токсина, встроили в гипервариабельный участок гена флагеллина Salmonella и полученную конструкцию ввели в дефектный по флагеллину штамм Salmonella. При этом было известно, что эпитоп, включающий 50 - 64 аминокислотные остатки субъединицы В холерного токсина, индуцирует выработку антител к интактному холерному токсину. Химерный флагеллин нормально функционировал, а эпитоп холерного токсина размещался на поверхности жгутиков. Иммунизация мышей с помощью интраперитонеальной инъекции примерно 5 - 106 живых или убитых формалином бактерий с модифицированным флагеллином индуцировала выработку большого количества антител как к пептиду ( 50 - 64 аминокислотным остаткам), так и к молекуле интактного холерного токсина. [10]
Именно такой подход использовали при создании противохолерной вакцины. Синтетический олигонуклеотид, кодирующий эпитоп субъединицы В холерного токсина, встроили в гипервариабельный участок гена флагеллина Salmonella и полученную конструкцию ввели в дефектный по флагеллину штамм Salmonella. При этом было известно, что эпитоп, включающий 50 - 64 аминокислотные остатки субъединицы В холерного токсина, индуцирует выработку антител к интактному холерному токсину. Химерный флагеллин нормально функционировал, а эпитоп холерного токсина размещался на поверхности жгутиков. Иммунизация мышей с помощью интраперитонеальной инъекции примерно 5 - 106 живых или убитых формалином бактерий с модифицированным флагеллином индуцировала выработку большого количества антител как к пептиду ( 50 - 64 аминокислотным остаткам), так и к молекуле интактного холерного токсина. [11]
Именно такой подход использовали при создании противохолерной вакцины. Синтетический олигонуклеотид, кодирующий эпитоп субъединицы В холерного токсина, встроили в гипервариабельный участок гена флагеллина Salmonella и полученную конструкцию ввели в дефектный по флагеллину штамм Salmonella. При этом было известно, что эпитоп, включающий 50 - 64 аминокислотные остатки субъединицы В холерного токсина, индуцирует выработку антител к интактному холерному токсину. Химерный флагеллин нормально функционировал, а эпитоп холерного токсина размещался на поверхности жгутиков. Иммунизация мышей с помощью интраперитонеальной инъекции примерно 5 - 106 живых или убитых формалином бактерий с модифицированным флагеллином индуцировала выработку большого количества антител как к пептиду ( 50 - 64 аминокислотным остаткам), так и к молекуле интактного холерного токсина. [12]
Во-вторых, программированной гибели клеток не обязательно предшествует снижение уровня АТФ. Более того, иногда программированная гибель наблюдается только в присутствии АТФ, что, по-видимому, связано с участием цАМФ в данном процессе. Особенно это характерно для клеток лимфоидного происхождения. Например, при инкубации клеток лимфомы с агентами, вызывающими увеличение фонда эндогенного цАМФ ( изопротеренол, ПГЕ) холерный токсин), наступают остановка роста и последующая гибель клеток [ Coffi - no Ph. В данном случае цАМФ выступает в роли индуктора, вызывающего дифференцировку и программированную гибель клеток. [13]
Привлекают внимание многочисленные радиобиологические эксперименты с кофеином, являющимся ингибитором фосфодиэстеразы. На Т - клетках почки человека показано, что кофеин ( 1 мМ) при добавлении в среду до и после облучения уменьшает индуцированную радиацией задержку митоза в стадии Ga. Подобным действием обладает МЭА, но при добавлении в среду только до облучения, менее эффективны теофиллин и теобромин. На синхронизированных культурах Physarum polycephalum, облученных в дозе 30 Гр, было продемонстрировано, что модификаторы уровня цАМФ по-разному влияют на задержку деления клеток: кофеин, теобромин и теофиллин уменьшают, дб-ц АМФ, холерный токсин, 1-метил - З - изобутилксантин и параверин не влияют, a RO-20-1724 увеличивает задержку деления. [14]
Грамотрицательная бактерия Vibrio cholerae вырабатывает белок-холерный токсин ( мол. Если не восполнять эти потери, то происходит тяжелое обезвоживание организма; без лечения болезнь часто приводит к смерти. Попавший в кишечник человека холерный токсин прочно связывается со специфическими участками на плазматической мембране эпителиальных клеток, выстилающих тонкий кишечник, и тем самым вызывает продолжительную ( измеряемую часами и днями) активацию аденилатциклазы. [15]
Страницы: 1 2