Транспозоны
Cтраница 3
Хотя, как мы убедились, многие свойства мобильных элементов и плазмид разумно рассматривать с точки зрения концепции эгоистичной ДНК, они играют существенную позитивную роль в жизни бактерий-хозяев, даже если не приносят непосредственной пользы. Дело в том, что IS-элементы, транспозоны и плазмиды служат важнейшими факторами генетической изменчивости и эволюции бактерий. [31]
В генетике бактерий все большее значение приобретает метод получения мутаций с помощью транс-позонов. Транспозоны ( Тп) представляют собой короткие двойные цепи ДНК, которые состоят из более чем 2000 пар оснований и обычно обусловливают устойчивость к одному антибиотику, в исключительных случаях-к нескольким, Транспозоны способны перепрыгивать из одного участка генома в другой, в частности из бактериальной хромосомы в плазмиду и обратно; таким образом, они могут включаться в различные участки генома ( см. разд. В случае внедрения транспозо-на в какой-либо структурный ген хромосомы нуклеотидная последовательность этого гена будет нарушена и генетическая информация не сможет транслироваться в функционально полноценный полипептид. [32]
Мигрирующие элементы могут встраиваться в разные участки бактериальной хромосомы или мигрировать с бактериальной хромосомы на плазмиду; их репликация осуществляется под контролем тех же механизмов, что и у соответствующей хромосомы или плазмиды. IS-элементы содержат информацию, необходимую только для их переноса внутри клетки, никаких выявляемых признаков в них не закодировано. Транспозоны устроены более сложно: в них включены некоторые гены, не имеющие отношения к процессу транспозиции. Известны транспозоны, содержащие гены устойчивости к антибиотикам, ионам тяжелых металлов и другим ингибиторам. Для переноса мигрирующих элементов между клетками нужен переносчик, которым могут быть определенные плазмиды или фаги. Встраивание мигрирующих элементов в бактериальную хромосому оказывает мутагенное действие, так как при этом происходит включение фрагмента ДНК, приводящее к изменению порядка расположения нуклеотидов в триплете и, как следствие этого, нарушению процесса транскрипции. [33]
Другой тип подвижных генетических элементов представляют транспозоны. Это участки генома, которые могут менять свое положение на хромосоме или же переходить с хромосомы на экстрахромосомные элементы. Транспозоны имеют по концам последовательности для включения ( IS - insertion sequence), а в середине может быть заключен любой ген, Перенос обслуживается специальным ферментом. Транспозоны могут быть перенесены на другое место на хромосоме, плазмиде, фаге. [34]
Транспозоны способны перемещаться по хромосоме. Их положение сказывается на экспрессии генов. Транспозоны могут существовать и вне хромосомы ( автономно), но неспособны к автономной репликации. [35]
Тп-элсмснты ( сложные перемещающиеся элементы, или транспозоны) принципиально отличаются от 18-элементов только тем, что содержат дополнит, структурные гены, не имеющие отношения к ф-ции транспозиции. Известно много транспозонов, в состав к-рых входят гены устойчивости к антибиотикам, тяжелым металлам и др. ядам. Существуют транспозоны, содержащие гены, к-рые кодируют токсины, а также свойственные данному организму ферменты. Как правило, Тп-элементы несут на концах целые или частично измененные 18-элементы, к-рые сообщают им способность перемещаться по геному и вызывать в нем те же изменения, что и своб. При этом 2 концевые 18-подобные терминальные последовательности в зависимости от типа транспозона могут иметь прямую или инвертир. Разные транспозоны часто содержат одинаковые терминальные последовательности нуклеотидов. [36]
Ясно, что описанный механизм транспозиции годится не для всех транспозонов. Например, составные транспозоны, состоящие из какого-либо гена, окруженного IS-элементами в прямой взаимной ориентации, по структуре неотличимы от коинтеграта ( ср. Далее, многие транспозоны образуют коинтеграты, отличающиеся исключительной стабильностью. [37]
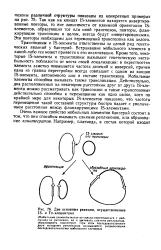 |
Две основные реакции, осуществляемые IS - и Тп-элементами. [38] |
Встраивание мобильного элемента в какой-либо ген может привести к его инактивации. Кроме того, некоторые IS-элементы и транспозоны вызывают генетическую нестабильность поблизости от места своей локализации: в окрестностях элемента заметно повышается частота делеций и инверсий, причем одна из границ перестройки всегда совпадает с одним из концов IS-элемента, автономного или в составе транспозона. Мобильные элементы способны вызывать также транслокации. Действительно, два расположенных на некотором расстоянии друг от друга IS-элемента можно рассматривать как транспозон и такие транспозоны действительно способны к перемещению как одно целое, хотя по крайней мере для некоторых IS-элементов показано, что частота перемещения такой составной структуры быстро падает с увеличением расстояния между фланкирующими IS-элементами. [39]
Этот элемент представлен в геноме 4 - 35 копиями, локализация к-рых отличается у разных штаммов. Копии Ту1 не полностью идентичны друг другу и составляют таким образом гетерог. В том случае, если две копии Ту1 заключают между собой клеточные гены, они перемещают и по геному, т.е. образуют истинные транспозоны. Включение Ту1 - подобных элементов в регуляторные зоны генов может вызывать не только инактивацию локусов, но и изменения механизма их регуляции, что, по-видимому, связано с присутствием в нуклеотидной последовательности Ту1 специфич. [40]
Другой тип подвижных генетических элементов представляют транспозоны. Это участки генома, которые могут менять свое положение на хромосоме или же переходить с хромосомы на экстрахромосомные элементы. Транспозоны имеют по концам последовательности для включения ( IS - insertion sequence), а в середине может быть заключен любой ген, Перенос обслуживается специальным ферментом. Транспозоны могут быть перенесены на другое место на хромосоме, плазмиде, фаге. [41]
Мак Клинток, которая в опытах с кукурузой показала, что гены могут перемещаться в пределах генома и влиять на механизмы экспрессии. В дальнейшем было установлено, что это явление характерно для многих эукариотических и прокариотических клеток. Транспозоны эукариотических клеток гораздо больше и включают в себя набор различных генов. Внутригеномное перемещение и встраивание транспозонов требует разрыва и последующего сращивания цепи ДНК. Репликация транспозона в одном сайте цепи, а затем перемещение и репликация в другом создают благоприятные возможности для дальнейших гомологичных рекомбинаций в клетке. [42]
Мигрирующие элементы могут встраиваться в разные участки бактериальной хромосомы или мигрировать с бактериальной хромосомы на плазмиду; их репликация осуществляется под контролем тех же механизмов, что и у соответствующей хромосомы или плазмиды. IS-элементы содержат информацию, необходимую только для их переноса внутри клетки, никаких выявляемых признаков в них не закодировано. Транспозоны устроены более сложно: в них включены некоторые гены, не имеющие отношения к процессу транспозиции. Известны транспозоны, содержащие гены устойчивости к антибиотикам, ионам тяжелых металлов и другим ингибиторам. Для переноса мигрирующих элементов между клетками нужен переносчик, которым могут быть определенные плазмиды или фаги. Встраивание мигрирующих элементов в бактериальную хромосому оказывает мутагенное действие, так как при этом происходит включение фрагмента ДНК, приводящее к изменению порядка расположения нуклеотидов в триплете и, как следствие этого, нарушению процесса транскрипции. [43]
Ключевым свойством бактериальных мобильных элементов, обеспечивающим их сохранение, является их способность перемещаться с репликона на репликон. Наличие у бактерий трансмиссивных и мобилизуемых плазмид позволяет транспозонам и IS-элементам не только переходить с плазмиды на плазмиду или из хромосомы на ллазмиду, но и путешествовать из клетки в клетку в составе плазмид. Таким путем мобильные элементы могут распространяться в бактериальных популяциях, даже если не приносят своим хозяевам никаких преимуществ. В этой связи следует упомянуть о явлении иммунности к транспозиции; многие транспозоны и IS-элементы значительно чаще перемещаются на новые репликоны, чем на новое место в составе того репликона, в котором они находятся. Молекулярный механизм этого свойства еще не выяснен, но очевидно, что оно способствует распространению мобильного элемента по максимальному количеству репликонов. [44]
Наличие у бактерий трансмиссивных и мобилизуемых плазмид позволяет транспозонам и IS-элементам не только переходить с плазмиды на плазмиду или из хромосомы на плазмиду, но и путешествовать из клетки в клетку в составе плазмид. Таким путем мобильные элементы могут распространяться в бактериальных популяциях, даже если не приносят своим хозяевам никаких преимуществ. В этой связи следует упомянуть о явлении иммунности к транспозиции; многие транспозоны и IS-элементы значительно чаще перемещаются на новые репликоны, чем на новое место в составе того репликона, в котором они находятся. Молекулярный механизм этого свойства еще не выяснен, но очевидно, что оно способствует распространению мобильного элемента по максимальному количеству репликонов. [45]
Страницы: 1 2 3 4