Транспорт - электрон
Cтраница 1
 |
Схематическое изображение бислойной мембраны с включенными в нее протеинами и транспортными комплексами. [1] |
Транспорт электронов и молекул через мембраны представляет другую область исследования, основанную на понимании структуры и организации мембран. [2]
Транспорт электронов по цепи электронного переноса п фотосинтезирующих системах, транспорт электронов по системе ферментов дыхательной цепи митохондрий, аллостерические свойства некоторых ферментов и многое другое, несомненно, указывает на большую эффективность переноса зарядов, энергии и информации на значительные расстояния в биосистемах. [3]
Тогда транспорт электронов по L становится возможным за счет небольшого количества тепловой энергии. [4]
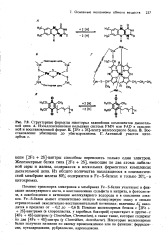
Помимо транспорта электронов в мембранах Fe-S - белки участвуют в фиксации молекулярного азота, в восстановлении сульфита и нитрита, в фотосинтезе, в освобождении и активации молекулярного водорода и в окислении алка-нов. Некоторые железосерные белки получили название по своему происхождению или по функции: ферредок-син, путидаредоксин, рубредоксин, адренодоксин. [6]
Нециклический же транспорт электрона от воды к НАДФ при фотосинтезе и сопряженный с ним синтез АТФ интенсивнее протекают при коротковолновом освещении ( ЛХ700 нм) и связаны с фотосистемой II. По мере повышения длины волны с 664 нм и дальше интенсивность нециклического фото-фосфорилирования сильно снижается и почти полностью прекращается при 708 - 714 нм. Циклическое же фотофосфорилирование при этих условиях усиливается, достигая максимума при 714 нм, после чего наблвдается небольшое снижение. [7]
Имеет место циклический транспорт электронов. [8]
На систему транспорта электронов оказывает влияние и О-хлорбензилхлорид. [9]
Основные пути транспорта электронов в ходе первичных процессов фотосинтеза показаны на рис. 12.14. Это известная Z-схема - результат исследований, в которых использовались методы импульсной спектро-фотометрии, а также искусственные доноры и акцепторы электронов и специфические ингибиторы. Она дает представление об окислительно-восстановительных потенциалах пигментов и переносчиков электронов и о последовательности их окисления и восстановления, но ничего не говорит о локализации этих компонентов в мембране. [10]
Псевдоциклический путь транспорта электрона и сопряженное с ним фотофосфорилирование также чувствительны к ингибиторам выделения кислорода. [11]
Важнейшие особенности транспорта электрона в РЦ - его высокая скорость и направленность в плотном мак-ромолекулярном комплексе, где исключены обычные диффузионные активам, механизмы хим. реакций в растворах по типу сталкивающихся частиц. [12]
Медиаторный механизм транспорта электрона достаточно широко используется для проведения электрохимических ферментативных реакций. В работе [45] описан электрохимический процесс с участием лактатдегидрогеназы, флавинмононуклео-тида и феназинметасульфата в качестве медиаторов; в работе ( 12 ] проанализирована электрохимическая ионизация водорода на угольных электродах, катализируемая гидрогеназой; в работе i [47] исследованы эффекты ускорения ионизации кислорода в системе пероксидаза - ионы металла - гидрохинон - пи-рографитовый электрод. [13]
Биологическое окисление и транспорт электронов по цепи дыхания тесно связаны с окислительным фосфорилированием, являющимся главным источником накопления свободной энергии в клетках в легко используемой форме - в виде богатых энергией фосфорных соединений, главным образом в АТФ. В окислительном цикле трикарбоновых кислот на каждую молекулу уксусной кислоты, окисленной до двуокиси углерода, образуется 8 протонов и 8 электронов, которые транспортируются по цепи дыхания и восстанавливают молекулярный кислород в воду. [14]
У бактерий хроматиум нециклический и циклический транспорт электронов осуществляется при участии одного и того же пигмента Р8эо, но реакционные центры ( Р и Р), в состав которых входит этот пигмент, по-видимому, пространственно ( физически) разделены и спектры действия этих разных фотосистем несколько отличаются. [15]
Страницы: 1 2 3 4