Гидродинамическое измерение
Cтраница 2
В обоих этих соотношениях s представляет собой коэффициент седиментации для предельного случая нулевой концентрации и с - концентрацию, выраженную в граммах в единице объема. Эффект концентрации играет значительно большую роль, чем в случае коэффициентов диффузии, поскольку последние зависят от отношения величины термодинамического члена к / [ ср. Экспериментальные значения коэффициентов седиментации всегда экстраполируются к нулевой концентрации по той же самой причине, что и при всех гидродинамических измерениях, а именно в связи с тем, что механические трактовки ( в данном случае для коэффициента трения) являются верными только в этих предельных условиях. [16]
Как видно из фиг. Два из них необходимы для ферментативной активности. Гидродинамические измерения показывают, что молекула пепсина обладает весьма компактной структурой, хотя в ней, по-видимому, имеется лишь небольшое число а-спиральных участков. [17]
Для некоторых белков, и в частности рибону-клеазы ( фермента, расщепляющего рибонуклеиновую кислоту), получены данные, позволяющие ответить на этот вопрос. Рибо-нуклеаза состоит из 124 аминокислотных остатков, последовательность которых известна. В нативном белке эти остатки, соединяясь попарно ( 26 - 84, 40 - 95, 58 - ПО, 65 - 72), образуют четыре дисульфидных мостика; существование этих мостиков затрудняет образование а-спирали. Как показывают гидродинамические измерения, молекула рибонуклеазы имеет почти сферическую форму, и, следовательно, цепь молекулы должна быть свернута, образуя третичную структуру, стабилизируемую взаимодействием боковых цепей. [18]
Если результаты гидродинамических измерений интерпретировать упрощенно, то можно прийти к выводу, что гидратацион-ные слои дают вклад в инерционность молекулы белка ( в растворе), а время жизни молекулы воды в этих слоях должно быть велико но сравнению с характеристическим временем движения белковых молекул. Например, если ориентационные времена релаксации больших белковых молекул, согласно, скажем, измерениям частотной зависимости диэлектрической проницаемости или вычислениям из закона Стокса, составляют 10 - 5 с, то можно ожидать, что продолжительность их жизни будет значительно больше. Однако такая точка зрения неправильна. Чтобы объяснить данные гидродинамических измерений, достаточно предположить, что при гидратации белка молекула воды находится в определенном положении. Если это условие удовлетворяется ( а для этого молекула воды должна резко изменить свой угловой момент инерции, как если бы она внедрялась в массу растворителя или же покидала его), то продолжительность жизни молекулы воды может быть меньше, чем время, которое характеризует движение белка и еще дает вклад в инерционность белковых молекул. На то, что это требование удовлетворяется, указывают рентгенографические данные, согласно которым многие молекулы воды в гидратационном слое находятся в определенных положениях. [19]
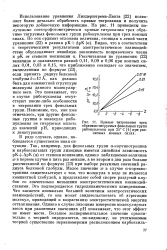 |
Кривые титрования трех обратимо титруемых фенольных групп рибонуклеазы при 25 С при различных ионных силах. [20] |
В ряде случаев, однако, наблюдается существенно иная ситуация. Так, например, для фенольных групп а-кортикотропина и карбоксильных групп лизоцима имеется линейная зависимость рК -) - ЛрК ( а) от степени ионизации, однако наблюдаемая величина w в первом случае в пять раз меньше, а во втором в два раза больше рассчитанной по формуле ( 23) при выборе разумных значений радиуса белковой глобулы. Малое значение w в случае а-кортикотропина свидетельствует о том, что молекула в воде не является компактной глобулой, а представляет собой скорее раздутый клубок со значительно меньшими электростатическими взаимодействиями. Это подтверждается гидродинамическими измерениями. Что касается аномально большой величины электростатических взаимодействий, то она может свидетельствовать об агрегации молекул, приводящей к возрастанию электростатической энергии отталкивания. Однако в случае лизоцима измерение молекулярных весов при различных значениях рН показывает, что этот эффект не имеет места. [21]
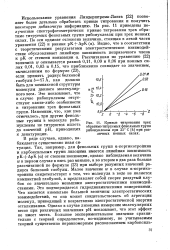 |
Кривые титрования трех обратимо титруемых фенольных групп. [22] |
В ряде случаев, однако, наблюдается существенно иная ситуация. Так, например, для фенольных групп а-кортикотропина и карбоксильных групп лизоцима имеется линейная зависимость рК -) - АрК ( а) от степе ни ионизации, однако наблюдаемая величина w в первом случае в пять раз меньше, а во втором в два раза больше рассчитанной по формуле ( 23) при выборе разумных значений радиуса белковой глобулы. Малое значение w в случае а-кортикотропина свидетельствует о том, что молекула в воде не является компактной глобулой, а представляет собой скорее раздутый клубок со значительно меньшими электростатическими взаимодействиями. Это подтверждается гидродинамическими измерениями. Что касается аномально большой величины электростатических взаимодействий, то она может свидетельствовать об агрегации молекул, приводящей к возрастанию электростатической энергии отталкивания. Однако в случае лизоцима измерение молекулярных весов при различных значениях рН показывает, что этот эффект не имеет места. [23]
В таких условиях коэффициент теплоотдачи следует определять с помощью широко известной аналогии Рейнольдса [22] в ее оригинальной или модифицированной форме. Эта аналогия позволяет связать трение и тепловой поток, используя стандартные безразмерные критерии. Затем с помощью гидродинамических измерений определяют параметры и трения, и теплообмена. Следовательно, данные по теплообмену нужно получить эмпирически и скоррелировать их при помощи аналитических методик. Разумеется, вопрос о том, в какой степени эти результаты применимы к нагревателю двигателя Стирлинга, остается открытым из-за отсутствия экспериментальных данных для таких условий течения. [24]
Нужно заметить, что эквивалентная гидродинамическая сфера представ ляет намного более простую концепцию, чем эквивалентная сфера, используемая для расчета исключенного объема. Это есть мера занятого объема, измененного межмолекулярными взаимодействиями. Когда силы притяжения становятся большими, как это имеет место в плохом растворителе, они играют доминирующую роль, и термодинамически исключенный объем может быть равным 0 или даже отрицательным. Гидродинамические измерения, однако, производятся ( или экстраполируются) в очень разбавленных растворах, в которых межмолекулярные взаимодействия вообще не играют роли. Гидродинамический объем всегда является мерой объема, занятого в растворе отдельной частицей. При расчете этого объема полимерная частица представляется до некоторой степени аналогичной гелю. Содержание воды в геле может достигать 99 %, однако эта жидкость оказывается неспособной к самостоятельному течению. [25]
Проблема равномерного подвода реагентов к слою катализатора не относится к категории новых. Традиционные способы решения этой проблемы предусматривают установку рассеивающих поток газораспределительных устройств в местах сосредоточенного ввода реагентов в аппарат, расположение выравнивающих поток решеток как в пространстве перед слоем, так и непосредственно на границе слоя. Естественно, что появляются энергозатраты на преодоление сопротивления газораспределительных решеток. Для уменьшения неоднородностей течения в прилегающих к слою областях увеличивают высоту этих областей, что приводит к дополнительному росту размеров аппарата. Экспериментальная проверка эффективности применяемых мер делается либо в отсутствие зернистого слоя, либо со слоем, однородность структуры которого не контролируется, что приводит к неоправданному увеличению гидравлического сопротивления или размеров аппарата. Для определения оптимальных способов борьбы с неоднородпостями, связанными с течением в прилегающих к слою областях, или же для оптимальной организации течения в этих областях необходимо экспериментировать на моделях со слоем, который должен иметь строго однородную структуру. Для построения математической модели, описывающей гидродинамику аппаратов с НЗС, полезно на двух-трех простых моделях аппаратов провести подробные гидродинамические измерения: определить линии тока, измерить как нормальную, так и касательную составляющие скорости газа на выходе из слоя, оценить характерную длину релаксации гидродинамических микроиеоднород-ностей ( струй) при вытекании из слоя. С помощью основанной на этих экспериментах математической модели можно просчитать десятки и сотни схем реакторов и найти для практического применения наиболее подходящие по своим технологическим характеристикам. [26]
При сравнении этих данных мы должны учитывать возможность того, что электронная микрофотография является обманчивой. Может быть, что замороженная и высушенная молекула, которую видно в электронный микроскоп, отличается от молекулы в растворе. Возможно также и то, что нити, которые соединяют шарики ( и которые плохо видны), простираются за пределы крайних шариков, так что истинная молекулярная длина превышает размер 475 А, измеренный между внешними кромками шариков. Сомнения относительно точности данных, получаемых при помощи электронного микроскопа, яляются, однако, неуместными в отношении главного обстоятельства, которое выявляется при сравнении двух результатов. Это обстоятельство состоит в том, что размеры, приведенные на рис. 37, являются абсолютно возможными и не несовместимыми с гидродинамическими данными. Поскольку уравнения для интерпретации гидродинамических данных являются ограниченными в том смысле, что они требуют использования модели вытянутого эллипсоида для всех жестких частиц, из этого следует, что мы можем вычислять по гидродинамическим данным только размеры эллипсоида, наиболее точно соответствующего действительной молекуле. Различие между лучшим эллипсоидом для фибриногена и формой, данной на рис. 37, должно рассматриваться как мера неточности, которая относится ко всем формам, выведенным из гидродинамических измерений, даже когда они комбинируются с данными по рассеянию света. [27]
Страницы: 1 2