Тэнфорд
Cтраница 2
Эти эффекты связываются [324] с электростатическим влиянием ( эффект поля) отрицательно заряженных атомов заместителей X, расположенных ближе к реакционному центру, чем положительные концы соответствующих диполей. Можно согласиться с авторами [324], что эти результаты не могут быть понятны с точки зрения модели передачи индукционного влияния через связи. Что же касается попытки количественной интерпретации в рамках модели Кирквуда - Вэстхэймера с использованием модификации Тэнфорда [7], то и в этом случае не удается добиться успеха без привлечения искусственных построений, касающихся вычисления значения эффективной диэлектрической проницаемости. [16]
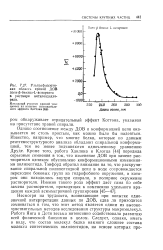 |
Ультрафиолетовая область кривой ДОВ поли-р-бензил - Ь - аспартата D растворе метилендихло-рида. [17] |
Однако соотношение между ДОВ и конформацией цепи оказывается не столь простым, как можно было бы надеяться. Известно, например, что многие белки, которые по данным рентгеноструктурного анализа обладают спиральной конформацией, тем не менее подчиняются одночленному уравнению Друде. Кроме того, работа Ханлона и Клотца [44] породила серьезные сомнения в том, что изменение ДОВ при замене растворителя целиком обусловлено конформационными изменениями полипептида. Тэнфорд обратил внимание на то, что раскручивание спиральной полипептидной цепи в воде сопровождается переходом боковых групп из гидрофобного окружения, з котором они находились внутри спирали, в полярное окружение растворителя. [18]
Благодаря большому электростатическому потенциалу, создаваемому заряженной макромолекулой в целом, она притягивает к себе низкомолекулярные противоионы из раствора, образуя как бы ионную шубу. Эти низкомолекулярные противоионы экранируют взаимодействие между заряженными группами макромолекулы, так что / ион и ДрК оказываются тем меньше, чем больше ионная сила раствора. Точный вид зависимости Ряоя и ДрК от заряда молекулы и ионной силы раствора определяется геометрией макромолекулы. В случае глобулярных белков обычно используют формулу Тэнфорда, основанную на предложенном Линдерштремом-Лангом рассмотрении компактной белковой глобулы как сферы с равномерно распределенным по поверхности зарядом. [19]
Благодаря большому электростатическому потенциалу, создаваемому заряженной макромолекулой в целом, она притягивает к себе низкомолекулярные противоионы из раствора, образуя как бы ионную шубу. Эти низкомолекулярные противоионы экранируют взаимодействие между заряженными группами макромолекулы, так что Рят и АрК оказываются тем меньше, чем больше ионная сила раствора. Точный вид зависимости FaOK и АрК от заряда молекулы и ионной силы раствора определяется геометрией макромолекулы. В случае глобулярных белков обычно используют формулу Тэнфорда, основанную на предложенном Линдерштремом-Лангом рассмотрении компактной белковой глобулы как сферы с равномерно распределенным по поверхности зарядом. [20]
Хорошо известно, что растворенное вещество может взаимодействовать с растворителем, но об этом часто забывают при описании величины оптического вращения вещества в растворе. В растворе могут происходить сложные молекулярные взаимодействия, конформа-ционные изменения и изменения ионных частиц; очевидно, такие изменения будут влиять на оптическое вращение, и в результате зависимости от природы растворителя и ( или) рН раствора будут наблюдаться различные величины вращения. Для многих амфотерных веществ, таких, как аминокислоты, знак оптического вращения изменяется при изменении рН раствора. Этого можно было ожидать, так как удельные вращения часто малы ( 100) и их кривые дисперсии оптического вращения пересекают ось нулевого вращения в области 400 - 500 нм. Таким образом, изменение молекулярных частиц, например переход цвиттер-иона в соль недиссоциированной кислоты и амина, должно сильно влиять на вращение. Тэнфорд [ 26] показал, что даже такое незначительное изменение растворителя, как переход от 40 % - ного к 60 % - ному водному диоксану, приводит к изменению знака вращения N-ацетил-ь - глутаминовой кислоты при D-линии натрия. [21]
Как и в случае дикарбоновых кислот, для получения хорошего согласования с экспериментом необходимо допустить, что все заряженные группы находятся внутри полости на расстоянии примерно 1 А от ее поверхности. Такая схема допускает значительное варьирование кривых титрования за счет различного расположения кислых групп. Этот результат очень важен, поскольку он дает возможность объяснить некоторые аномалии кривых титрования, исходя из чисто электростатических представлений без введения вспомогательных постулатов ( таких, например, как водородная связь), даже если они и кажутся очевидными. Энергия электростатического взаимодействия минимальна при равномерном распределении зарядов, максимальна при резко неравномерном их распределении и имеет промежуточное значение, если заряды распределены беспорядочно. Так, например, скопление карбоксильных групп на небольшом участке приводит к возрастанию величины со. В одной из моделей Тэнфорда область минимума энергии электростатического взаимодействия не находится в изоэлектрической точке. Это дает возможность, по крайней мере качественно, объяснить тот опытный факт, что область максимальной стабильности некоторых белков не совпадает с изоэлектрической точкой. Следует помнить, что теория Тэнфорда - Кирквуда была разработана только для сферической модели. Между тем молекулы многих белков и нуклеиновые кислоты не являются сферическими, а потому эта теория к ним неприложима. Математический анализ следующей по сложности эллипсоидальной модели молекулы сопряжен с огромными трудностями. [22]
Как и в случае дикарбоновых кислот, для получения хорошего согласования с экспериментом необходимо допустить, что все заряженные группы находятся внутри полости на расстоянии примерно 1 А от ее поверхности. Такая схема допускает значительное варьирование кривых титрования за счет различного расположения кислых групп. Этот результат очень важен, поскольку он дает возможность объяснить некоторые аномалии кривых титрования, исходя из чисто электростатических представлений без введения вспомогательных постулатов ( таких, например, как водородная связь), даже если они и кажутся очевидными. Энергия электростатического взаимодействия минимальна при равномерном распределении зарядов, максимальна при резко неравномерном их распределении и имеет промежуточное значение, если заряды распределены беспорядочно. Так, например, скопление карбоксильных групп на небольшом участке приводит к возрастанию величины со. В одной из моделей Тэнфорда область минимума энергии электростатического взаимодействия не находится в изоэлектрической точке. Это дает возможность, по крайней мере качественно, объяснить тот опытный факт, что область максимальной стабильности некоторых белков не совпадает с изоэлектрической точкой. Следует помнить, что теория Тэнфорда - Кирквуда была разработана только для сферической модели. Между тем молекулы многих белков и нуклеиновые кислоты не являются сферическими, а потому эта теория к ним неприложима. Математический анализ следующей по сложности эллипсоидальной модели молекулы сопряжен с огромными трудностями. [23]
Страницы: 1 2