Изофермент
Cтраница 2

Один из изоферментов, катализирующих превращение аспартата в аспар-тилфосфат, может аллостерически инги-бироваться двумя различными модуляторами-лизином и изолейцином, действие которых более чем аддитивно. Это еще один пример согласованного ингиби-рования ( разд. [17]

Все пять изоферментов содержат в разных соотношениях полипептидные цепи двух видов, различающиеся по аминокислотному составу и последовательности аминокислот. А-цепи, называемые также М - це-пями ( от англ, muscle - мышца), кодируются двумя разными генами. Изофор-ма лактатдегидрогеназы, преобладающая в скелетной мышце, содержит четыре А-цепи, а изоформа, преобладающая в мышечной ткани сердца, состоит из четырех В-цепей. [19]
Почему клетки продуцируют изоферменты. [20]
Возможно, существуют изоферменты двух типов. [21]
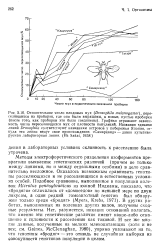
Методы электрофоретического разделения изоферментов превратили выявление генетических различий ( причем ие только между линиями, но и между отдельными особям-и) в дело сравнительно несложное. [23]
В отличие от изоферментов ЛДГ изоферменты ААП определяются в различных органах не в виде полного спектра ( 5 изоферментов), а чаще как один изофермент. Так, изофермент АА: представлен главным образом в ткани печени, ААП2 - в поджелудочной железе, ААП3 - в почках, ААП4 и ААП5 - в различных отделах стенки кишки. При повреждении ткани почек изофермент ААП3 обнаруживается в крови и моче, что является специфическим признаком поражения почечной ткани. [24]
В тканях позвоночных обнаружены изоферменты фосфорилазы, фосфоглюкомутазы, гексозоизомеразы, гексокиназы, фосфофруктокиназы, альдолазы, триозофосфатдегидрогеназы. Из этого ясно, что многие ( вероятно, все) гликолитиче-ские ферменты в мышце представлены изоферментами, более или менее специфичными для мышечной гкани. Детальные данные об их приспособленности для функционирования в микросреде мышечных клеток имеются не дли всех этих ферментов, но относительно некоторых получены вполне убедительные сведения; к их числу относится фосфофруктокиназа, которая, так же как фосфоглицераткиназа и пируваткиназа, катализирует реакции, сопровождающиеся значительным уменьшением свободной энергии: для ФФК AG - 3 4 ккал / моль, для ФГК - 4 5, а для ПК - 7 5 ккал / моль. [25]
В и С обозначают соответствующие изоферменты карбоангидразы эритроцитов человека. [26]
Таким образом, наличие изоферментов позволяет варьированием соотношения аллелей изменять суммарную каталитическую активность клетки и точнее регулировать интенсивность метаболических процессов. Возможность изменения соотношения аллелей появляется только у полиплоидов, а создание растения с наибольшим приближением к максимуму активности по всем ферментам и с наиболее точной регуляцией скоростей реакций и есть создание организма с оптимальной плоид - ностью ядра. Так как большинство ферментов состоит из нескольких субъединиц, то рассмотренная связь имеет, по всей видимости, большое значение. [27]
Аминокислотный состав этих пяти изоферментов также характеризуется постепенным изменением в пределах между двумя крайними формами. Иммунологические свойства и ферментативная активность обнаруживают аналогичные плавные переходы. [28]
Наличие в эритроцитах двух изоферментов карбоангидразы с различной первичной структурой является, по-видимому, отличительной особенностью приматов. [29]
В отличие от изоферментов ЛДГ изоферменты ААП определяются в различных органах не в виде полного спектра ( 5 изоферментов), а чаще как один изофермент. Так, изофермент АА: представлен главным образом в ткани печени, ААП2 - в поджелудочной железе, ААП3 - в почках, ААП4 и ААП5 - в различных отделах стенки кишки. При повреждении ткани почек изофермент ААП3 обнаруживается в крови и моче, что является специфическим признаком поражения почечной ткани. [30]
Страницы: 1 2 3 4