Изучение - кинетика - гидролиз
Cтраница 2

Недавно мы совместно с О. Г. Сердюк [51, 52] при изучении кинетики гидролиза фенил - и хлорфенолгликозидов получили данные, хорошо соответствующие конформационным представлениям схемы Эдварда и в свою очередь являющиеся новым фактическим материалом, подтверждающим ее. По нашим данным, фенил и пара-хлорфенилгликозиды гидролизуются медленнее, чем соответствующие ксилозиды, вследствие наличия у Сб-гликозидов тяжелого заместителя СНаОН, который при образовании формы полукресла должен выйти из экваториального положения и несколько приблизиться к аксиальному. С другой стороны, гидроксил у С4, находящийся у га-лактозидов в аксиальном положении, при образовании формы кресла будет переходить в положение, близкое к экваториальному, и этим облегчать гидролиз галактозидов по сравнению с гликозидами. Более медленный гидролиз изучавшихся нами п-хлорфенилгликози-дов по сравнению с таковым соответствующих фенилгликозидов может быть объяснен понижением электронной плотности в бензольном ядре агликона и говорит в пользу присоединения протона к кислородному атому замещенного полуацетального гидроксила, а не к кислородному мостику пиранозидного кольца. [17]
К аналогичному выводу приводит рассмотрение табл. 6.5, в которой сравниваются полученные при изучении кинетики гидролиза диарилметилгалогенидов в водном ацетоне значения констант а ( ос. Так, низкой скорости гидролиза соответствует малая величина константы а. Большая величина а в случае 4-фенилдифенилметилхлорида свидетельствует, по-видимому, о том, что высокая поляризуемость сообщает кар-бониевому иону дополнительную устойчивость и, тем самым, большую селективность тю отношению к одному из конкурирующих нуклеофильных агентов, но не влияет на переходное состояние, приводящее к образованию этого иона. [18]
Смете и Де Лекер [54] использовали сополимеры метилметак-рилата с метакриловой кислотой, полученные радикальной сополи-меризацией, для изучения кинетики гидролиза полиметилметакри-лата, однако набор констант k0, ki и kz им определить не удалось, что обусловлено, наряду с другими причинами, указанным ограничением применимости продуктов сополимеризации в качестве полимерных моделей ( напомним, что стереохимические эффекты вносят существенный вклад в кинетику гидролиза полиметилметак-рилата - см. гл. [19]
Этого недостатка лишен другой способ определения коэффициента распределения, упомянутый в монографии Гельфериха55 и успешно примененный Тар - тарелли5б 57 при изучении кинетики гидролиза эфи-ров низших алифатических спиртов. [20]
Тем не менее, поскольку метод Ван-Сляйка стандартизован до мельчайших деталей, он позволяет получать ценные сравнительные данные и, следовательно, является удобным средством изучения кинетики гидролиза. [21]
Исследование кинетики процесса гидролиза целлюлозы имеет важнейшее значение для выяснения условий проведения процесса гидролиза целлюлозосодержащих материалов в производственных условиях и для дальнейшего уточнения общих вопросов строения целлюлозы. При изучении кинетики гидролиза целлюлозы применяют различные методы. В последние годы, в связи с усиленным вниманием к этой проблеме были предложены новые методы исследования процесса гидролиза. [22]
В заключение интересно очень кратко рассмотреть влияние растворителя на скорость процесса нуклеофильного замещения. И, действительно, при изучении кинетики гидролиза бромистых алкилов, растворенных в муравьиной кислоте ( содержащей 0 5 % воды), которая, по сравнению с другими растворителями, характеризуется большой диэлектрической постоянной ( стр. [23]
Например, этому уравнению подчиняются экспериментальные данные работы ( Sodetz, Castellino, 1972), в которой исследовалась предстационарная кинетика действия плазминов. Аналогичные данные получены в работе Бендера и соавторов при изучении кинетики гидролиза л-нитрофенилацетата под действием а-химотрип-сина. [24]
Растворы этих солей постепенно выделяют водород. При подкислении выделение водорода усиливается. Изучение кинетики гидролиза декаборана показывает, что реакция идет по первому порядку. [25]
АВК дают продукты кислотного расщепления: фторированные кислоты, нефторированные метилкетоны и амины. При продолжительном выдерживании АВК в нейтральных водно-спиртовых средах изменений не происходит. Изучение кинетики гидролиза АВК показало, что константы скорости гидролиза в кислой среде изменяются в пределах четырех порядков, в зависимости от природы заместителей. Наличие фторалкильной группы у енаминного реакционного центра увеличивает скорость гидролиза. [26]
Интерес к гидролизу целлюлозы проявляется в течение многих лет и относится к проблемам аналитического и препаративного характера. Исследование процесса гидролиза целлюлозы представляет интерес с двух точек зрения. Во-первых, изучение кинетики гидролиза может быть принципиально использовано для получения информации о наличии в макромолекуле целлюлозы аномальных элементарных звеньев, отличающихся от обычных a - D-глю-копиранозных звеньев, связанных ( 1 - - 4) - р-связью. Во-вторых, процесс гидролиза целлюлозы в разбавленных кислотах всегда осложняется тем, что целлюлоза нерастворима в гидролизующей среде. [27]
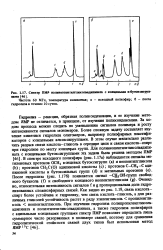 |
Спектр ПМ полинеопентилгликольадишшата с концевыми н-бутоксигруп-цами. [28] |
Гидролиз - реакция, обратная поликонденсации, и ее изучение методом ЯМР не отличается, в принципе, от изучения поликонденсации. Более сложную задачу составляет изучение кинетики гидролиза олигомеров, например полиэфирных пластификаторов с концевыми алкоксигруппами. В этом случае желательно различить разрыв связи кислота-гликоль в середине цепи и связи кислота-спирт при гидролизе по месту концев ых групп. [29]
Функциональные группы активного центра химотрипсина идентифицированы с помощью необратимых ингибиторов. Остаток Ser-195 был модифицирован диизопропилфторфосфатом и фенил-метилсульфофторидом, а остаток His-57 - г4 - тозил - Ь - фенилала-нил-хлорметилкетоном ( см. рис. 89 и с. Двухстадийность процесса химотрипсинового гидролиза была обнаружена при изучении кинетики гидролиза п-нитрофени л ацетата. [30]
Страницы: 1 2 3