Кристаллический фермент
Cтраница 2
Недавно исследованы рибонуклеазы, имеющие даже большую специфичность, чем основной компонент кристаллического фермента из поджелудочной железы. Конечным продуктом гидролиза дрожжевой рибонуклеиновой кислоты является гуанозин-3 - фосфат ( около 40 % общего содержания гуанина) и олигонуклеотиды с гуанозин-3 - фосфатом на конце. Как и в случае панкреатической рибонуклеазы, стадия трансфосфорилирования обратима [169, 170] и фермент катализирует как полимеризацию гуанозин-2 3 -циклофосфата до олиго-и, возможно, полигуаниловой кислот, так и трансфосфорилирова-ние 5 -гидроксильной группы нуклеозидов ( с образованием дину-клеозидфосфатов, таких, как гуанилил-3: 5 -уридин и гуанилил-3: 5 -аденозин), нуклеозид - З - фосфатов и нуклеозид-2 3 -цикло-фосфатов. [16]
С помощью аналитического ультрацентрифугирования было показано, что единственный пик, полученный при седиментации кристаллического фермента, распадался на два при добавлении стеарилкофермента А; при добавлении щавелевоуксусной кислоты диссоциация принимала обратимый характер. Очевидно, что концентрация щавелевоуксусной кислоты в ткани может играть важную роль в регуляции действия цитрат-синтазы не только потому, что она стабилизирует фермент, но также и потому, что она является его субстра гом. Так, при высокой интенсивности глкжонеогснеза концентрация щавелевоуксусной кислот i понижается в результате быстрого превращения в фосфоенолпируват. В этих условш-х печень компенсирует понижение скорости ф нкциони-рования цикла трикарбоновых кислот повышением скорости окислительных реакции, не входящих в цикл, а именно превращая жирные кислоты в ацетоацетат, который затеи переносится к различным тканям н окисляется. [17]
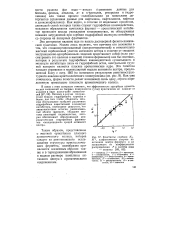 |
Константы сорбции / Сь М 1, алифатических спиртов на активном центре фермента и их зависимость от коэффициентов распределения Р в системе вода - октанол. [18] |
Таким образом, представление о жесткой ориентации плоского ароматического кольца, которое следует из рентгеновского исследования структуры кристаллического фермента, своеобразно проявляется косвенным образом также и в термодинамике образования в водном растворе комплекса активного центра с ароматическими соединениями. [19]
Представление об активном центре химотрипсина как структуре, состоящей из пространственно и функционально разделенных участков ( сорбционного и каталитического), в литературе сложилось раньше [102], чем появились рентгеновские данные о структуре кристаллического фермента. Использованные подходы могут оказаться весьма полезными при изучении структуры и функций активного центра других ферментов. [20]
 |
Механизм действия. [21] |
Глицерофосфатдегидрогеназная активность была обнаружена Эйлером и его сотрудниками в мышцах крыс. Кристаллический фермент был получен Барановским из мышц кролика. Высокой активностью он обладает в летательных мышцах насекомых, активность его в скелетных мышцах позвоночных животных намного выше, чем в сердечной мышце и почках, низкая активность в мозге. [22]
Выделение кристаллических ферментов способствовало пониманию природы их высокой специфичности, так как их белковая природа и данные о специфичности белковых веществ позволили строить гипотезы относительно особенностей их строения и неповторимости их функций в зависимости от деталей структуры. [23]
Ферменты, которые удалось выделить в кристаллическом состоянии, обладают очень высокой удельной активностью. Растворы кристаллических ферментов способны проявлять каталитические свойства при таких незначительных концентрациях фермента, при которых белковая природа фермента не может быть обнаружена обычными химическими реакциями на белок. [24]
Как оказалось, ферменты, приготовленные по Нортропу, являются прекрасно выраженными кристаллами белка, не оставляющими сомнения в их однородности. Впрочем, эти кристаллические ферменты мало похожи на обычные кристаллы минеральных веществ или органических соединений; они содержат значительное количество воды и солей и могут существовать только в насыщенном маточном растворе. При попытках высушивания они разрушаются. [25]
Как оказалось, ферменты, приготовленные по Нортропу, являются вполне однородными кристаллами белка. Впрочем, эти кристаллические ферменты мало похожи на обычные кристаллы минеральных веществ или органических соединений; они содержат значительное количество воды и солей и могут существовать только в насыщенном маточном растворе. При попытках высушивания они разрушаются. [26]
Вескими доказательствами белковой природы фермента являются его получение в чистом виде и выделение в форме кристаллов белка. К настоящему времени получено более 1000 кристаллических ферментов. [27]
По существу, все полученные до настоящего времени кристаллические ферменты оказались либо простыми, либо сложными белками ( протеидами) с определенной простетической группой. [28]
Условия растворимости ферментов сходны во всех отношениях с таковыми у глобулинов. Если бы не были известны необычайные каталитические свойства чистых кристаллических ферментов, то их можно было бы отнести, по-видимому, к обычным классам белков или протеидов. Подобно всем белкам, ферменты являются специфическими антигенами: при введении в кровь животного они вызывают образование антител. [29]
Эти результаты ( см. рис. 42) интересно сопоставить с данными по структуре кристаллического химотрипсина. Согласно Стейтцу и др. [66], гидрофобная полость в кристаллическом ферменте ( см. рис. 9) действительно узка ( 3 5 - 4 А) и длина ее не превышает 10 А. Таким образом, рентгеновские исследования структуры кристаллического фермента [66] и кинетические исследования его субстратной специфичности в водном растворе [21] привели к взаимно согласующимся данным о геометрии активного центра химотрипсина. [30]
Страницы: 1 2 3