Актиновый филамент
Cтраница 1
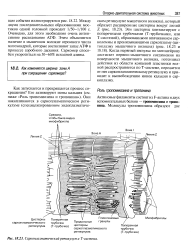 |
Саркоплазматический ретикулум и Т - система. [1] |
Актиновые филаменты состоят из F-актина и двух вспомогательных белков - тропомиозина и тропонина. [2]
Для передвижения конуса роста необходимы актиновые филаменты. Если в культуральную среду добавить цитохалазин В, предотвращающий полимеризацию актина, движение микропиишков конуса роста прекращается и аксон перестает продвигаться вперед. Тем не менее конус роста остается прикрепленным к субстрату, и нейрит сохраняет свою прежнюю длину. Если же в культуральную среду добавить колхицин, разрушающий микротрубочки, нейрит будет втягиваться обратно в тело клетки. В то же время из проксимальных участков нейрита, которые до того были гладкими и прямыми, будут развиваться новые микрошипики и даже новые конусы роста. Поэтому было высказано предположение, что микротрубочки служат для стабилизации удлиняющегося нейрита и для ограничения участков, где может проявляться активность конусов роста, подобно тому как в фибробласте микротрубочки, по-видимому, определяют локализацию мембранной складчатости ( см. разд. [3]
На каждой молекуле актина, входящей в актиновый филамент, имеются участки, комплементарные определенным участкам на головках молекул миозина и способные взаимодействовать с ними с образованием слабой связи ( см. разд. Комплементарность может быть нарушена в результате изменений в структуре взаимодействующих участков, причем это изменение может быть вызвано каким-либо источником энергии. При стимуляции сокращения мышцы комплементарные участки начинают соединяться. [4]

Наши сведения о структуре миозина растений пока довольно скудны, однако пучки актиновых филаментов найдены в самых разных растительных клетках, в том числе и в тех, что формируют волоски. Тот факт, что в большинстве клеток высших растений цитоплазма может двигаться во многих направлениях ( в отличие от однонаправленного тока, характерного для гигантских клеток водорослей), позволяет думать, что соседние пучки актиновых филаментов здесь могут иметь противоположную полярность. [6]

Клетки высших растений содержат все те внутриклеточные компартменты, которые ранее были описаны для животных клеток-это цитозоль, аппарат Гольдки, эндоплазматический ретикулум, ядро, митохондрии, пероксисомы и лизосомы. Кроме того, растительные клетки обладают цитоскелетом, состоящим из актиновых филаментов и микротрубочек, которые неотличимы от соответствующих структур в клетках животных. Однако в клетках растений имеются два дополнительных мембранных компартмента-вакуоли и пластиды. Последние два типа органелл связаны с неподвижным образом жизни растений. [8]

Электронно-микроскопические исследования показали, что эти фибриллы представляют собой пучки актиновых филаментов, ориентированных одинаковым образом. Полярность актиновых филаментов такова, что перемещение вдоль них других-миозиновых-филаментов могло бы создать наблюдаемый в клетке направленный ток цитоплазмы ( разд. Таким образом, не исключено, что органеллы в движущейся цитоплазме сцеплены с актиновыми филаментами, но не прямо, а через молекулы миозина, которые, используя энергию гидролиза АТР, скользят вдоль актиновых филаментов, увлекая за собой органеллы. [10]

Наши сведения о структуре миозина растений пока довольно скудны, однако пучки актиновых филаментов найдены в самых разных растительных клетках, в том числе и в тех, что формируют волоски. Тот факт, что в большинстве клеток высших растений цитоплазма может двигаться во многих направлениях ( в отличие от однонаправленного тока, характерного для гигантских клеток водорослей), позволяет думать, что соседние пучки актиновых филаментов здесь могут иметь противоположную полярность. [12]

Электронно-микроскопические исследования показали, что эти фибриллы представляют собой пучки актиновых филаментов, ориентированных одинаковым образом. Полярность актиновых филаментов такова, что перемещение вдоль них других-миозиновых-филаментов могло бы создать наблюдаемый в клетке направленный ток цитоплазмы ( разд. Таким образом, не исключено, что органеллы в движущейся цитоплазме сцеплены с актиновыми филаментами, но не прямо, а через молекулы миозина, которые, используя энергию гидролиза АТР, скользят вдоль актиновых филаментов, увлекая за собой органеллы. [14]

Страницы: 1 2