Форма - хлорофилл
Cтраница 2
Спектр живого листа зависит от одновременного присутствия разных форм хлорофилла а в разных соотношениях. Разнообразие форм хлорофилла с длинноволновым поглощением наблюдается у растений только in vivo. При растворении в органических растворителях все формы хлорофилла переходят в одну с максимумом поглощения ъ 660 - 670 нм. [16]
Зеленые пигменты растений - хлорофиллы, сенсибилизирующие процессы фотосинтеза, - также являются производными порфина, в которых атомы азота пиррольных групп связаны с магнием. Существуют несколько форм хлорофиллов, различающихся между собой только некоторыми радикалами, замещающими водородные атомы в р-положении пиррольных колец. Основным компонентом фотохимической системы высших растений и водорослей является хлорофилла. В фото-синтезирующих бактериях, в синезеленых водорослях и-в других организмах содержатся другие разновидности хлорофиллов и родственных им систем. [17]
Для того чтобы исследовать это взаимодействие, были поставлены опыты, в которых в отсутствие воздуха в трубку вводили раствор хлорофилла в пиридине с аскорбиновой кислотой, в отвод трубки - раствор окислителя-сафранина, рибофлавина, дифосфопиридин-нуклеотида, либо других соединений. Ори освещении трубки быстро образовывалась красная фотопос-становлеиная форма хлорофилла; затем в темноте из отвода приливали к фотопродукту раствор окислителя и следили за ходом реакции по величине коэффициента погашения в максимуме поглощения хлорофилла и его фотовосстановленной формы. Этот опыт, таким образом, дает возможность осуществить сенсибилизированный процесс раздольно по стадиям - световой и темповой. [18]
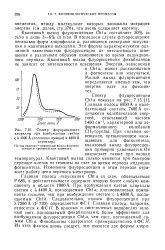 |
Спектр флуоресценции. [19] |
Квантовый выход флуоресценции Chi а составляет 30 % in vitro и лишь 3 - 6 % in vivo. В последнем случае флуоресцируют несколько форм хлорофилла а, разнящиеся спектрами испускания и квантовыми выходами. Эти формы характеризуются различной зависимостью флуоресценции от интенсивности освещения и различным образом изменяются во время индукционного периода. Квантовый выход флуоресценции фикобилинов не зависит от интенсивности освещения. [20]
Если красный цвет крови обусловлен наличием в ней тема, то зеленые тона растительного мира созданы хлорофиллом. В листьях растений присутствуют несколько форм хлорофилла. Преобладающими являются хлорофилл а 6.83 и хлорофилл b 6.84. Как видно из этих формул, хлорофиллы - производные хлориновой циклической системы. [21]
До сих пор неясно, что собой представляет реакционной центр. Вероятно, это одна из наиболее длинноволновых форм хлорофилла а, которая очень эффективно расходует энергию электронного возбуждения на осуществление фотохимической реакции. Скорость фотохимической окислительно-восстановительной реакции превышает скорость расходования энергии возбуждения на люминесценцию на два порядка величин у бактерий и почти на три порядка величин у высших растений. [22]
Спектр поглощения СЫ а имеет сложную полосу в красной области. Это заставляет думать о нескольких формах хлорофилла. [23]
Другая форма поглощает только более коротковолновый свет или возбуждается путем резонансной миграции энергии от дополнительного пигмента. Каждый из этих процессов вызывается главным образом одной из двух форм хлорофилла а. Эффективный фотосинтез происходит лишь тогда, когда обе формы хлорофилла а возбуждаются одновременно. [24]
Система II является вспомогательной пигментной системой. Она состоит примерно из 100 молекул Хл а 670, представляющего собой флуоресцирующую форму хлорофилла а, поглощающую в более коротковолновой области, чем Хл а системы I. [25]
Две фотохимические системы, которые можно обозначить ФС) и ФС2, взаимодействуют таким образом, что электроны системы ФС2 используются системой ФСь Система ФС ] представлена хлорофиллом а и другими пигментами, поглощающими красный дальний свет. Система ФС2 использует набор пигментов ( фикобилины, хлорофилл Ъ и одну форму хлорофилла а с максимумом поглощения при 670 ммк), которые поглощают свет более короткой волны. [26]
Световая энергия, воспринимаемая большим числом ( около 200) молекул хлорофилла, передается на единый активный центр. Поскольку расстояние между молекулами хлорофилла около 1 8 нм, для процесса передачи энергии синглетной формой хлорофилла за время жизни возбужденного состояния может осуществиться около 300 актов передачи энергии. При соотношении числа активных центров и числа воспринимающих пигментов 1 / 200 активные центры всегда получают достаточное количество энергии. Порядок расположения пигментов еще окончательно не выяснен. Обычно хлорофилл в биологических системах связан с белками, образуя с ними пигмент-белковые комплексы. При обработке пигментов поверхностно-активными веществами происходит сдвиг в длинах волн поглощаемого света. Этот факт сразу наводит на мысль об использовании полимерных матриц для моделирования и регулирования процессов описываемого типа. [27]
Вызываемое светом изменение поглощения происходит не только при зеленении этиолированных листьев. Путем изучения разностных спектров Кок [193-195] показал, что в водорослях и хлоропластах присутствует какой-то пигмент ( Р700) с максимумом поглощения около 700 нм, который, вероятно, представляет собой одну из форм хлорофилла а. По-видимому - при освещении дальним красным светом или при вспышках белого света высокой интенсивности ( ртутная дуговая лампа) этот пигмент быстро выцветает или разрушается, так как его поглощение быстро падает. [28]
 |
Фотосопротпв-1 - светочув-слой, 2 - ме-таллич. электроды, 3 - диэлектрич. подложка. [29] |
Миграция энергии между молекулами хлорофилла может идти на уровне возбужденных синглетных или тринлетных состояний, в форме экситона, электрона или дырки. Экспериментально установлена фотопроводимость твердых пленок хлорофилла и его аналогов. В результате миграции энергии возбуждение достигает наиболее низко расположенных форм хлорофилла, связанных с активными центрами, в к-рых происходит первичный фотохимич. Эффективная миграция энергии идет в упорядоченных агрегированных структурах, в к-рых взаимодействуют близко расположенные молекулы пигментов. [30]
Страницы: 1 2 3