Фосфорилаз
Cтраница 2
Ингибирует мышечную фосфорилазу, а также захват глюкозы эритроцитами. Механизм воздействия на эритроциты не совсем ясен. [16]
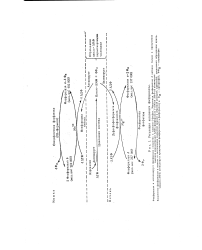 |
Регуляция активности фосфорилазы. [17] |
АМФ; фосфорилазу Ь печени АМФ не активирует. [18]
Определение активности крахмальной фосфорилазы. В основу метода положен принцип уменьшения неорганического и увеличения органического фосфора в реакционной смеси в процессе фосфоролити-ческого расщепления крахмала. [19]
В отличие от фосфорилазы b фос-форилаза а не активируется AMP; поэтому фосфорилазу а называют иногда АМР-независимой формой, а фосфорилазу b - AMP-зависимой. Итак, есть два механизма регуляции, которым подчиняется гликоген-фосфорилаза скелетной мышцы: 1) ковалентная модификация посредством фосфоршшрования или де-фосфорилирования остатков серина в активном центре фермента и 2) аллостери-ческая регуляция фосфо рилазы b путем нековалентного связывания с AMP или АТР. [20]
Определение активности образующейся фосфорилазы а. Активность фермента измеряют по обратной реакции ( синтезу гликогена), сопровождающейся выделением неорганического фосфата. К малеат-ному буферу ( 0 1 М), рН 6 5, содержащему 0 1 М глюкозо-1 - фосфат - 2 % - ный гликоген, добавляют равный объем раствора, полученного после киназной реакции. Реакцию проводят 5 мин при 30 С. Останавливают реакцию добавлением реактивов для определения неорганического фосфата. Количество образовавшегося фосфата рассчитывают по калибровочному графику. [21]
В скелетной мускулатуре фосфорилаза находится в двух формах: b и а. Активность фосфорилазы b можно определить только в присутствии АМФ; фосфорилаза а активна в отсутствие АМФ. Для обеих форм фермента АМФ является положительным аллостерическим эффектором. Молекула фосфорилазы b представляет собой димер, фосфорилазы а - тетрамер. Молекулярная масса субъединицы фермента равна 97400 Да. Обе формы фермента могут находиться в состоянии равновесия между димерными и тетрамерными молекулами. На переход димеров в тетрамеры и обратно оказывают влияние компоненты ферментативной реакции, активаторы, ингибиторы, а также рН, ионная сила раствора, температура и др. Наиболее активными являются димеры обеих форм. [22]
В этой реакции фосфорилаза а превращается в фосфорилазу Ь, гораздо менее активно катализирующую распад гликогена. Таким образом, активная форма гликогенфосфорилазы превращается в относительно неактивную форму в результате расщепления двух ковалент-ных связей между остатками фосфорной кислоты и двумя специфическими остатками серина в молекуле фермента. [23]
Сахаров, катализируемые фосфорилазами; 2) реакции переноса с участием нуклеотидов, содержащих сахара; 3) реакции переноса без участия фосфорных эфиров, катализируемые трансгликозилазами нефосфоролитич. В последнюю группу входят реакции ферментативного гидролиза Сахаров. [24]
Фосфорилазу а получают из фосфорилазы Ъ, освобожденной от АМФ ( с. Реакцию катализирует киназа фосфорилазы в присутствии Mg-АТФ. [25]
Гири и сотрудники обнаруживают фосфорилазу на поверхности агара, содержащего глюкозо-1 - фосфат, и после инкубации обрабатывают агар раствором йода. [26]
Этот процесс, регулируемый фосфорилазами, в сущности аналогичен процессу гидролиза, только вместо воды в нем участвует фосфорная кислота, поэтому он и называется фосфоролизом. [27]
Хотя здесь речь шла о мышечных фосфорилазах, основные принципы действия этого фермента относятся и к печеночной фосфорилазе. Последняя также существует в двух формах - активной и малоактивной; и их взаимопревращениями регулируется фосфоролиз в печени. Однако печеночная фосфорилаза отличается от мышечной как по строению, так механизму регуляции, что вполне понятно исходя из различий динамики обмена гликогена в мышцах и печени. [28]
Активная протеин-киназа катализирует фосфорилирование ферментов - фосфорилазы и гликогенсинтазы. Образуется активная фосфорилированная форма фосфорилазы а и неактивная фосфорилированная форма гликогенсинтазы D; такие изменения обеспечивают фосфоролити-ческий распад гликогена. [29]

Страницы: 1 2 3 4