Фосфорилирование
Cтраница 2
Фосфорилирование происходит в трех узловых пунктах - в участке цепи НАД. [16]
Фосфорилирование и де-фосфорилирование модифицирует их свойства. Способность НГБ стимулировать синтез РНК в бесклеточной системе зависит от состояния их фосфорилирования. Гипотеза состоит в том, что ген включается присоединением негистонового белка к специфическому участку ДНК, репрессированному гистоном. НГБ фосфорилируется и приобретает отрицательные заряды. [17]
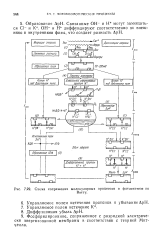 |
Схема сопряжения молекулярных процессов в фотосинтезе по. [18] |
Фосфорилирование, сопряженное с разрядкой электрически энергизованной мембраны в-соответствии с теорией Митчелла. [19]
Фосфорилирование, возможно, является частью процесса инактивации, который следует за световой активацией родопсина. [20]
Фосфорилирование, сопряженное с циклическим потоком электронов, получило название циклического фотофос-форилирования. Соответственно, нециклическим фо-тофосфорилированием называют синтез АТФ, сопряженный с нециклическим электронным транспортом. [21]
Фосфорилирование, сопряженное с переносом электронов от субстратов в темновых окислительных реакциях, получило название окислительного фосфорилирования. [22]
Фосфорилирование - 1) замещение атома водорода в молекуле органического соединения остатками фосфорных кислот; 2) образование фосфатного производного биомолекулы обычно за счет ферментативного переноса фосфатной группы по АТР. [23]
Фосфорилирование в этих условиях алкилгалогенидов приводит к смеси органических фосфинов и дифосфинов. Невысокий выход продуктов реакции и ее низкая селективность объясняются, вероятно, недостаточной эффективностью расщепления связей Р - Р в макромолекуле красного фосфора и образованием в результате полифосфид-анионов, а также фосфидов меньшей степени полимеризации и большей степени замещения. [24]
Фосфорилирование протекает в диффузионной области с энергией активации - для хлорида олова ( IV) ЗпСЦ - 34 0 кДж / моль. [25]
Фосфорилирование, фосфорилирование в цикле трикарбоновых к-т) и в биомембранах ( окислит, фосфорилирование, фо-тофосфорилирование) принципиально различаются по механизму преобразования энергии. В растворимых системах синтез АТФ, как правило, сопряжен с окислением альдегидных групп ( фос-фоглицериновый альдегид, янтарный полуальдегид и др.) пиридиннуклеотидами или флавопротеидами. Обычно альдегиды самопроизвольно взаимодействуют с Н - группой фермента или кофермента, происходит окисление комплекса, образуются макроэргич. [26]
Фосфорилирование может происходить только в тех участках, в которых сопряженная реакция ( реакция 5.3) протекает с уменьшением свободной энергии. [27]
Фосфорилирование, сопряженное с переносом электронов. Предположение о такого рода фосфорилировании у сульфатредуцирующих бактерий основано на данных о наличии цитохромов и железосерных белков в плазматических мембранах, а также о высоком выходе энергии. [28]
Фосфорилирование самого креатина в мышечной ткани с образованием фосфокреатина в конечном счете осуществляется за счет энергии двух первых процессов. [29]
Фосфорилирование dCDP с образованием dCTP ( стадия л, рис. 14 - 29) завершает биосинтез первого из пиримидиновых предшественников ДНК. Уридиннуклеотиды получаются двумя путями. Интересно, что ДНК-по-лимеразы способны включать это соединение в полинуклеотиды. [30]
Страницы: 1 2 3 4