Меченый фрагмент
Cтраница 1
Меченые фрагменты разделяют так же, как и при секвенировании ДНК по Максаму - Гилберту. [1]
Гибридизацию меченых фрагментов ДНК с ДНК-агаром ведут в стеклянных флаконах с притертой пробкой в термостате при 60 в 4XSSC в течение 16 - 18 часов. [2]
Такую смесь меченых фрагментов олигонуклеотида снова подвергают двумерному фракционированию электрофорезом на ацетил-целлюлозе и гомохроматографией на ДЭАЭ-целлюлозе, как описано выше, а затем проводят авторадиографию. Однако теперь вместо разбросанных по всей пластинке пятен фингерпринта получается зигзагообразная цепочка следующих друг за другом пятен ( рис. 173), анализ расположения которых дает информацию о последовательности нуклеотидов в олигонуклеотиде. [3]
Полученная смесь меченых фрагментов ДНК подвергается электрофорезу в денатурирующем вертикальном полиакриламид-ном геле. [4]
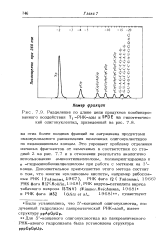 |
Разделение по длине цепи продуктов комбинированного воздействия Т - РНК-азы и SPDE на гипотетический олигонуклеотид, приведенный на. [5] |
Это упрощает проблему отделения меченых фрагментов от немеченых в соответствии со стадией 2 на рис. 7.7 и в отношении результата аналогично использованию аминоэтилцеллюлозы, полиакрилгидразида и п - гидразинобензилцеллюлозы при работе с метками на 3 -конце. [6]
На схеме мы видим, что меченые фрагменты, полученные при выщеплении остатков G, движутся со скоростью, соответствующей олигонуклеотидам длиной в 5 и 9 нуклеотидных звеньев; таким образом, остатки 6 и 10 в исходном олигонуклеотиде должны быть G. В нижней части рис. 1 дана нуклеотидная последовательность, установленная с помощью этой простой процедуры и некоторых рассуждений. Этим методом часто быстрее чем за 2 дня может быть определена нуклеотидная последовательность олигонуклеотидов, содержащих 200 и даже большее число остатков. [7]
Для выделения этих белков используют различные методы обнаружения их специфических комплексов с небольшими мечеными фрагментами ДНК, содержащими мотивы. [8]
Для выделения этих белков используют различные методы обнаружения их специфических комплексов с небольшими мечеными фрагментами ДНК, содержащими мотивы. [9]
Аналогичные возможности заключает в себе и метод малоуглового рассеяния медленных нейтронов, однако его использование требует применения меченых фрагментов сетки, распределение и размеры которых являются предметом изучения. Обычно для этой цели используют дейтериро-ванные соединения. [10]
Несвязанные меченые фрагменты ДНК отмывают примерно 20 - 50 мл 4XSSC до исчезновения радиоактивности в промывных водах. Затем из агара извлекают ДНК-ДНК-гибриды путем обработки 0 01X X SSC при 75 и собирают фракции по 5 мл. [11]
 |
Фрагменты частичного гидролиза 5 -меченого олигонуклеотида фос-фодиэстера 1пй змеиного яда. [12] |
В методе блуждающего пятна эти две задачи решаются одновременно. Смесь меченых фрагментов подвергается двумерному разделению. [13]
После иммобилизации мембранные фильтры промывают раствором 6SSC, просушивают в течение 12 час. Гибридизацию с мечеными фрагментами реперной ДНК проводят в бюксах при температуре инкубации, ниже температурит плавления ДНК на 25, в течение 16 - 18 час. [14]
В результате обнаружения явления необратимого ингибиро-вания холинэстераз органическими фосфорными соединениями и того факта, что фосфорильный остаток вида ( RO) 2P O соединяется с ферментом, возникла новая возможность изучения структуры активной поверхности. Если фосфорильный радикал пометить изотопом, то последующее разложение молекулы инги-бированного фермента должно дать меченый фрагмент, представляющий собой исходный компонент, который прореагировал с ингибитором. В такого рода исследованиях фосфорильный радикал неизменно получается связанным с р-гидроксилом серина. Естественно возникает вопрос, представляет ли выделенный меченый фрагмент первичный комплекс фермент-ингибитор, образующийся на активной поверхности, или же он является вторичным продуктом, получающимся в результате перефосфори-лирования во время гидролиза. [15]
Страницы: 1 2