Ацентрический фрагмент
Cтраница 2
Асимметричные межхромосомные обмены, касаются ли они целых хромосом или отдельных хроматид, вероятно, нежизнеспособны, поскольку они ведут к образованию ацентрических фрагментов. Помимо утери фрагмента, приводящей к нехватке, существование дицентрических хромосом, которые иногда образуют мостики, вызывает механические затруднения в процессе деления. [16]
В первой анафазе в результате подобного перекреста появляется дицентрическая хроматида 1 - 2 - 3 - 1, которая растягивается между двумя центромерами, ацентрический фрагмент 4 - 2 - 3 - 4 и две нормальные хроматиды 1 - 2 - 3 - 4 и / 3 - 2 - 4, которые соединены с дицентрической хроматидой центромерными участками. Во время второго мейотического деления эти хроматиды отделяются от дицентрической хроматиды. [17]
В первой анафазе эти центромеры направляются к противоположным полюсам, и поэтому хроматида тянется в виде моста через всю клетку от одного полюса к другому. Ацентрический фрагмент располагается вблизи моста. [18]
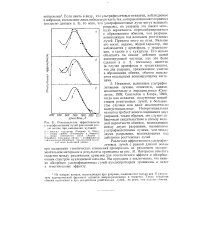 |
Относительная эффективность ультрафиолетовых лучей различной длины волны при вызывании мутаций. [19] |
Из четырех концов, возникающих при разрыве, соединяются только два. В результате ацентрический фрагмент остается неприкрепленным и теряется. Такие неполные обмены возникают и при воздействии рентгеновых лучей, но реже, чем полные обмены. [20]
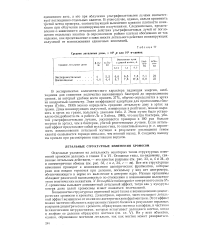 |
Средние летальные дозы, X Ю3 р или 103 в-единиц. [21] |
Все эти структурные изменения приводят к возникновению ацентрических фрагментов, которые рано или поздно теряются при делении, поскольку у них нет центромера, обеспечивающего в норме их включение в дочернее ядро. Разные организмы обладают различной выносливостью по отношению к возникающим вследствие этого генетическим нехваткам. У Drosophila melanogaster потеря всего лишь 5 % Х - хромосомы вызывает доминантный летальный эффект, тогда как у кукурузы потеря даже целой хромосомы может оказаться нелетальной. [22]
Большинство структурных изменений ведет также к возникновению дицент-рических хромосом и хроматид. Дицентрики, вероятно, часто вызывают летальный эффект независимо от их связи с ацентрическими фрагментами. Этот эффект можно частично объяснить нарушением генного баланса в результате неравных разрывов дицентрических хромосом, образующих мостики в анафазе, и частично механическими затруднениями, которые испытывает делящаяся клетка, когда в анафазе ее деления образуются мостики ( см. гл. Не у всех объектов, однако, образование мостиков летально, так как мостик может разорваться. [23]
 |
Число клеток с моносошшми в культурах ФЧ, обработанных стрептолизином-0, на фоне введения в культуру аутогенных Т - лимфо-цитов ( широкие столбики - до введения, узкие - после введения. [24] |
В анализируемых культурах после введения Т - лимфоци-тов и воздействия стрептолизина-0 наблюдалось достоверное изменение соотношения числа клеток с различными типами цитогенетических нарушений. Введение неиммунных аутогенных Т - лимфоцитов резко уменьшает число клеток с анеуплоидным хромосомным набором, не влияя на их частоту с обменами, точечными ацентрическими фрагментами и полиплоидным хромосомным набором. [25]
При делении клетки хромосомы удваиваются и затем расходятся так, что каждая новая клетка получает один из этих наборов хромосом. Хемостерилизаторы могут реагировать с одной из хроматид или со всей хромосомой. Когда имеется больше одного разрыва, может происходить воссоединение с образованием дицент-рических хромосом и ацентрических фрагментов. В дицентрическои хромосоме обе воссоединившиеся хромосомы сохраняют центромеры двух исходных хромосом, а ацентрический фрагмент не содержит центромеры. [26]
 |
Проявление гетерозиса в FI при скрещивании мутанта с исходным сортом. / - исходный сорт. 2 - мутант с редуцированными листочками. 3 - гибрид / V. [27] |
Инверсия ( поворот участка хромосомы на 180) приводит к образованию петли в профазе мейоза на стадии пахинемы. В результате кроссинговера внутри такой петли могут возникать при наличии инверсии, не захватывающей район центромеры, дицентрические ( с двумя центромерами) хромосомы, образующие мосты при расхождении хромосом в анафазе I ( А I, а в некоторых случаях и в Л II), сопровождаемые ацентрическими фрагментами. При наличии таких мостов в ряде клеток одного пыльника можно говорить о наличии у ( мутанта инверсии. [28]
При делении клетки хромосомы удваиваются и затем расходятся так, что каждая новая клетка получает один из этих наборов хромосом. Хемостерилизаторы могут реагировать с одной из хроматид или со всей хромосомой. Когда имеется больше одного разрыва, может происходить воссоединение с образованием дицент-рических хромосом и ацентрических фрагментов. В дицентрическои хромосоме обе воссоединившиеся хромосомы сохраняют центромеры двух исходных хромосом, а ацентрический фрагмент не содержит центромеры. [29]
С помощью электронного микроскопа установлено, что микроядра не имеют полной ядерной оболочки и в них отсутствует периферический конденсированный хроматин. Количество пор на ядерной оболочке не зависит от размеров микроядра. Как правило, микроядра, локализованные в одной клетке, отличаются по размерам в зависимости от содержания в них ДНК, при этом ДНК способна к репликации и время репликации в различных микроядрах не совпадает. Образование крупных микроядер ( диаметр 2 - 3 мк) тесно связано с геномными нарушениями, в то время как уровень клеток с мелкими микроядрами коррелирует с частотой нарушений в структуре хромосом. Клетки с ацентрическими фрагментами предшествуют появлению клеток с микроядрами. Однако не всегда отмечается подобная закономерность. [30]
Страницы: 1 2