Ингибирующая функция
Cтраница 2
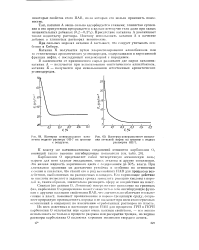 |
Изотерма поверхностного натяжения водного раствора ОП-7 на границе с воздухом.| Изотерма поверхностного натяжения атовской нефти на границе с водным раствором ОП-7. [16] |
Логинова) поверхностное натяжение па границах фаз, карбозолин О одновременно может совместить в себе ингибирующие функции с другими важными свойствами ПАН, что значительно облегчает его вытеснение в пласт, повышает проникновение в норово-трещшшую среду, сохраняет природную проницаемость породы и не вызывает при всем этом серьезных осложнении в операциях по извлечению отработанных растворов из пласта. [17]
Нами был также рассмотрен вопрос о возможном объяснении наблюдаемых явлений тормозящим действием стеарата кобальта, который использовался в качестве катализатора при окислении бутана. Десятикратное увеличение концентрации катализатора не изменяет характера протекания процесса окисления бутана, что свидетельствует об отсутствии ингибирующей функции катализатора в наших условиях. [18]
Нами был также рассмотрен вопрос о возможном объяснении наблюдаемых явлений тормозящим действием стеарата кобальта, который использовался в качестве катализатора при окислении бутана. Десятикратное увеличение концентрации катализатора не изменяет характера протекания процесса окисления бутана, чтс свидетельствует об отсутствии ингибирующей функции катализа-тор а в наших условиях. [19]
Коррозия оборудования технологических цепей добычи нефти остается одной из наиболее актуальных проблем в отрасли. Активно протекающие коррозионные процессы в системах поддержания пластового давления ставят ближайшей целью создание эффективных химических средств защиты от коррозии - реагентов с ингибирующими функциями. [20]
 |
Зависимость периода индукции ги в реакции окисления н. декана от концентрации введенного стеарата меди. [21] |
Таким образом, катализаторы переменной валентности ( соли кобальта, марганца и меди) не только инициируют, но и обрывают цепи. В зависимости от условий может преобладать одна из этих функций катализатора, в результате чего он выступает то в роли инициирующего агента, то в роли ингибитора. Подробное количественное изучение ингибирующей функции катализатора заслуживает самого серьезного внимания. [22]
Таким образом, катализаторы переменной валентности - соли кобальта, марганца и меди - не только инициируют, но и обрывают цепи в окисляющемся циклогексане. В зависимости от условий, которые существуют в реакции, может преобладать одна из этих функций катализатора и он выступает то в роли инициирующего агента, то в роли ингибитора. Подробное количественное изучение ингибирующей функции катализатора небходимо. Такого рода исследования только начинают ставиться, и мы не располагаем данными по всем этим вопросам. [23]
 |
Зависимость периода индукции ги в реакции окисления н. декана от концентрации введенного стеарата меди. [24] |
Таким образом, катализаторы переменной валентности ( соли кобальта, марганца и меди) не только инициируют, но и обрывают цепи. В зависимости от условий может преобладать одна из этих функций катализатора, в результате чего он выступает то в роли инициирующего агента, то в роли ингибитора. Подробное количественное изучение ингибирующей функции катализатора заслуживает самого серьезного внимания. [25]
В развившейся реакции инициирование цепей в результате взаимодействия восстановленной формы катализатора с гидроперекисью преобладает над обрывом цепей на молекулах катализатора. Поэтому в развившемся окислении ингибирующая функция катализатора практически не проявляется. Ее можно обнаружить, измеряя лишь порядок скорости реакции по концентрации катализатора. [26]
 |
Карта комплементации для 5 % - ного уровня отсечения между девятью изученными S-аллелями. [27] |
Если принять, что с разными аллелями связана различная продуктивность, т.е. образуется разное число полипептидов в единицу времени ( это утверждение несколько модифицирует гипотезу Сэмпсона [23]), то у гегерозигот концентрация одного гомок1ультнмера будет еще более низкой и изменятся относительные концентрации гибридных белков. Если имеются какие-то пороговые концентрации, при которых белок вьшолняет свою функцию, то для разных аллелей время, необходимое для достижения пороговой концентрации, неодинаково. У тетраплоидов концентрация гомомультимерного белка может оказаться ниже порогового уровня, и у этих белков ингибирующая функция будет отсутствовать. Хорошим подтверждением этому является случай, когда при наличии одной и той же пары аллелей может проявляться как конкурентное взаимоотношение, так и отношение доминирования. Подводя итог всему вышеизложенному, можно предложить следующий механизм совместимости у тетраплоидов. В результате образования гибридного белка в диплоидном пыльцевом зерне концентрация гомомультимерных белков, а следовательно и их активность уменьшаются. Если гибридные молекулы, соответствующие данному генотипу, не имеют ингибирующей активности или она очень мала, а активность гомомультимеров несколько ниже пороговой, то тетраплоид будет самосовместим. [28]
Из сказанного не следует, что в развившейся реакции окисления катализаторы переменной валентности не обрывают цепи. Обрыв их на катализаторе, по-видимому, имеет место и в развившейся реакции, однако в этих условиях инициирующая функция катализатора намного превосходит его ингибирующую способность. Кроме того, при высокой концентрации свободных радикалов в развившейся реакции скорость их квадратичного обрыва велика и по сравнению с ней линейный обрыв цепей на молекулах катализатора незначителен. Ингибирующая функция катализатора отчетливо проявляется, как мы видели, в начале реакции, когда катализатор не инициирует цепей. [29]
ЛТК в отличие от ТК являются ингибиторами автолизинов. Механизм их действия основан на том, что встроенная гидрофобным концом в цитоплазматическую мембрану ЛТК, соединенная через холин с автолизином, удерживает его в мембране, не допуская фермент к субстрату - ПГ. При действии деацилаз ЛТК, отщепляющих ацильный гидрофобный остаток молекулы, они теряют способность удерживаться в мембране, что приводит к потере их ингибирующих свойств. Помимо ЛТК ингибирующими функциями обладают некоторые мембранные липиды, такие как кардиолипин или дипальми-тоилфосфатидилглицерин. Поэтому мембранные липиды и механизмы их метаболических превращений играют важную роль в регуляции автолитических процессов и автолизе клетки. [30]
Страницы: 1 2 3