Хейдон
Cтраница 1
Хейдон и Филлипс отметили при этом необходимость использования тщательно очищенной воды, особенно в опытах с анионными ПАВ. [1]
Хейдон и Филлипс [11] определили межфазное натяжение растворов додецилсульфата натрия и додецилтриметиламмонийбромида на поверхности раздела углеводород - вода и отметили, что явление старения зависит от чистоты используемой воды. [2]
Хейдон и Филлипс отметили при этом необходимость использования тщательно очищенной воды, особенно в опытах с анионными ПАВ. [3]
Хейдон ( 1958) вычислил величину работы неоднородной адсорбции. Наблюдения за каплей воды, взвешенно: в толуоле с 4 % ацетона показывают, что эта капля испытывает беспорядочные пульсации. Частота этой пульсации уменьшается с течением времени, и движение исчезает, когда ацетон распределяется между толуолом и водой в соответствии с коэффициентами распределения. Количество энергии, затраченной в процессе пульсации, может быть определено по амплитуде или иной характеристике пульсации. Поглощенная энергия может быть определена по изменению физико-химических свойств системы. [4]
Хейдон ( 1958) вычислил величину работы неоднородной адсорбции. Наблюдения за каплей воды, взвешенной в толуоле с 4 % ацетона показывают, что эта капля испытывает беспорядочные пульсации. Частота этой пульсации уменьшается с течением времени, и движение исчезает, когда ацетон распределяется между толуолом и водой в соответствии с коэффициентами распределения. Количество энергии, затраченной в процессе пульсации, может быть определено по амплитуде или иной характеристике пульсации. Поглощенная энергия может быть определена по изменению физико-химических свойств системы. [5]
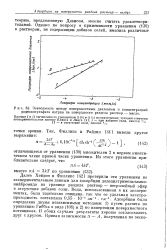 |
Зависимость между поверхностным давлением и концентрацией додецилсульфата натрия на поверхности раздела раствор - масло. [6] |
Далее Хейдон и Филлипс [42] проверили эти уравнения по экспериментальным данным для адсорбции додецилтриметиламмо-нийбромида на границе раздела раствор - петролейный эфир в отсутствие добавок соли. Адсорбция была определена двумя независимыми методами: 1) путем расчета по уравнению Гиббса в виде ( 108) и 2) по измерениям поверхностного потенциала. В обоих случаях результаты хорошо согласовались друг с другом. [7]
Бисвас и Хейдон получили двумерные релаксационные кривые предела текучести пленки методом Тачибана и Инокучи ( 1953) и выразили их в форме реологической модели Максвелла - Войгта, определив таким образом цифровые данные для коэффициентов эластичного сдвига и вязкости. В действительности они нашли, что эти две величины тесно связаны. Это объясняется образованием молекулами протеина сетчатых структур. [8]
Сривастава и Хейдон ( 1964), исследуя высокостабильные к коалесценции эмульсии ( 1 % - ный петролейный эфир, диспергированный в 0 1 о-ном бычьем сывороточном альбумине 0 01 М раствор КС1), смогли измерить скорость флокуляции. Оказалось, что около изо-электрической точки протеина флокуляция происходила в течение нескольких часов; при других значениях эмульсии становились полностью диспергированными или некоторое время слабо агрегировали. [9]
Бисвас и Хейдон получили двумерные релаксационные кривые предела текучести пленки методом Тачибана и Инокучи ( 1953) и выразили их в форме реологической модели Максвелла - Войгта, определив таким образом цифровые данные для коэффициентов эластичного сдвига и вязкости. В действительности они нашли, что эти две величины тесно связаны. Это объясняется образованием молекулами протеина сетчатых структур. [10]
Сривастава и Хейдон ( 1964), исследуя высокостабильные к коалесценции эмульсии ( 1 % - ный петролейный эфир, диспергированный в 0 1 % - ном бычьем сывороточном альбумине 0 01 М раствор КС ]), смогли измерить скорость флокуляции. Оказалось, что около изо-электрической точки протеина флокуляция происходила в течение нескольких часов; при других значениях эмульсии становились полностью диспергированными или некоторое время слабо агрегировали. [11]
Бисвас и Хейдон получили двумерные релаксационные кривые предела текучести пленки методом Тачибана и Инокучи ( 1953) и выразили их в форме реологической модели Максвелла - Войгта, определив таким образом цифровые данные для коэффициентов эластичного сдвига и вязкости. В действительности они нашли, что эти две величины тесно связаны. Это объясняется образованием молекулами протеина сетчатых структур. [12]
Сривастава и Хейдон ( 1964), исследуя высокостабильные к коалесценции эмульсии ( 1 % - ный петролейный эфир, диспергированный в 0 1 % - ном бычьем сывороточном альбумине 0 01 М раствор КС1), смогли измерить скорость флокуляции. Оказалось, что около изо-электрической точки протеина флокуляция происходила в течение нескольких часов; при других значениях эмульсии становились полностью диспергированными или некоторое время слабо агрегировали. [13]
Для систем В / М исследования на липидных мембранах ( Хейдон: и Тейлор, 1966; Зонтаг, Нетзель и Кларе, 1966) показали, что даль-недействующее отталкивание отсутствует, но маслорастворимое ПАВ обусловливает эластичность пленки В / М / В, которая затем быстро утончается до бимолекулярной структуры. Этот бимолекулярный слой удерживает некоторое количество растворителя и является жидким; вандерваальсово сжатие уравновешивается осмотическими силами. [14]
Для систем В / М исследования на липидных мембранах ( Хейдон и Тейлор, 1966; Зонтаг, Нетзель и Кларе, 1966) показали, что даль-недействующее отталкивание отсутствует, но маслорастворимое ПАВ обусловливает эластичность пленки В / М / В, которая затем быстро утончается до бимолекулярной структуры. Этот бимолекулярный слой удерживает некоторое количество растворителя и является жидким; вандерваальсово сжатие уравновешивается осмотическими силами. [15]
Страницы: 1 2