Природная целлюлоза
Cтраница 2
Волокна природной целлюлозы могут быть окрашены крч-сителями различных типов. [17]
В природной целлюлозе не удается показать наличие редуцирующих групп из-за их очень малого содержания. Технические целлюлозы всегда более или менее разрушены и в зависимости от типа разрушения содержат большее или меньшее число редуцирующих групп. При гидролитическом расщеплении целлюлозы в месте разрыва глюкозидной связи возникает одна нередуцирующая и одна редуцирующая группа. При окислительном разрушении целлюлозы могут возникать самые разнообразные группы и в том числе редуцирующие. Определение числа редуцирующих групп имеет важное значение для характеристики средней длины цепных молекул целлюлозы, а также для характеристики степени разрушения и чистоты технических целлюлоз. Многие исследователи [1, 2, 3, 4] связывают с числом редуцирующих групп способность беленых целлюлоз к пожелтению. [18]
Кристаллическую модификацию природной целлюлозы называют целлюлозой I, а модификацию гидратцеллюлозы - целлюлозой II. Кроме того, у гидрат-целлюлозы ослабляются водородные связи и увеличивается содержание аморфной части. [19]
 |
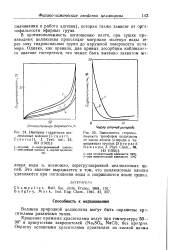
Параллельный наклонный спектр кристаллитов целлюлозы II рами, ориентированных в двух направлениях. Электрический вектор и ось цепи направлены перпендикулярно щели. [20] |
В мембранах природной целлюлозы плоскость 101 преимущественно параллельна поверхности пленки. [21]
Многие виды природной целлюлозы отличаются низкой степенью кристалличности, поэтому дифракция рентгеновских лучей не является идеальным методом для установления преобладающего расположения цепей. Значительное увеличение при повороте интенсивности полос валентных колебаний О - Н - групп при частоте 3350 и 3275 см - указывает на высокую степень ориентации в одной плоскости для регенерированной целлюлозы и на расположение плоскости, в которой в основном находятся водородные связи, перпендикулярно поверхности пленки. Поскольку многие целлюлозные мембраны и полученные в производственных условиях пленки обладают лишь одноплоскостной ориентацией, а не осевой, наклонный дихроизм представляет собой ценный метод для исследования морфологической структуры. [22]
Изменение кристалличности природной целлюлозы при высушивании, установленное методом дейтерирования [50, 51], дает ряд ценных сведений о механизме образования структуры этой целлюлозы. [23]
Для препаратов природной целлюлозы, отличающихся более плотной упаковкой, частицы, которые уже нельзя считать независимыми, дают рефлексы брэгговского типа; таким образом можно отделить данные о размерах частиц от сведений об их распределении. Отнесение непрерывного экваториального малоуглового рассеяния в сухой целлюлозе к микропустотам [21], по-видимому, требует нового объяснения результатов некоторых прежних исследований. Рассеяние рентгеновских лучей под малыми углами используется для изучения конформа-ций макромолекул некоторых биополимеров в растворе. [24]
При набухании природной целлюлозы в воде изменения рентгенограммы не наблюдается и, следовательно, образования гидрата не происходит. [25]
В препаратах природной целлюлозы и гидратцеллюлозы в отдельных микрообластях степень упорядоченности и ориентации макромолекул или их звеньев различна, что приводит к различной скорости диффузии реагентов и, в результате, к различной скорости реакций в отдельных участках волокна. Все реальные препараты целлюлозы структурно неоднородны. [26]
Как для природной целлюлозы, так и для гид-ратцеллюлозы, и в частности для вискозного шелка, характерно более быстрое протекание реакции в начальной стадии и замедление ее в последующих стадиях процесса гидролиза. [27]
Обычно препараты природной целлюлозы со степенью полимеризации ниже 200 представляют собой порошки. [28]
Рентгенограммы волокон природной целлюлозы показывают, что кристаллиты имеют преимущественную ориентацию вдоль оси волокна. [29]
Однако у природной целлюлозы рентгенографически длинные периоды обнаружить еще не удалось. Гесс и Киссиг 86 связывают их с периодическим чередованием кристаллических и аморфных областей. У синтетических волокон длинные периоды связывают со складками молекул, причем длина периода соответствует одной складке. Тоннесен и Эллефсен и особенно Долмеч полагают, что складчатая структура характерна также для целлюлозы. Хозе-манн 87 рассматривает длинные периоды, как следствие паракристалличе-ской структуры решетки, которая состоит из статистически изогнутых слоев. Петерлин 86 объясняет возникновение длинных периодов термодинамическими затруднениями. На экваторе рентгенограммы также был обнаружен максимум, который в набухшем состоянии равен 70 - 110 А, а в сухом состоянии - 45 - 55 А. Последние данные совпадают с результатами исследований под малыми углами рассеяния. [30]
Страницы: 1 2 3 4