Активный центр - химотрипсин
Cтраница 2
Относительно низкий уровень собственной реакционной способности, который обнаруживает ферментный, нуклеофил в свободном хи-мотрипсине, имеет, по-видимому, важное биологическое значение. Дело в том, что такое свойство активного центра химотрипсина значительно ограничивает возможность неспецифических реакций при физиологических условиях. [16]
Однако при добавлении внешнего бензоат-иона ( в роли третьего компонента активного центра химотрипсина выступает карбоксилат-ион) скорость гидролиза этой модельной системы только в четыре раза ниже скорости ферментативного деацилирования ( в предположении, что фактор перевода межмолекулярных реакций IBO внутримолекулярный режим равен 10 М, см. разд. [17]
Оказалось, что этот реагент селективно алкилирует остаток ги-стидина ( His-57), расположенный в активном центре химотрипсина. [18]
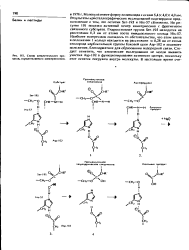 |
Схема каталитического процесса, осуществляемого химотрипсином. [19] |
Молекула имеет форму эллипсоида с осями 5 4X4 0X4 0 нм. Результаты кристаллографических исследований подтвердили предположение о том, что остатки Ser-195 и His-57 сближены. На рисунке 101 показан активный центр химотрипсина с фрагментом связанного субстрата. Гидроксильная группа Ser-195 находится на расстоянии 0 3 нм от атома азота имидазольного кольца His-57. Наиболее интересным оказалось то обстоятельство, что атом азота в положении I кольца находится на расстоянии 0 28 нм от атома кислорода карбоксильной группы боковой цепи Asp-102 и занимает положение, благоприятное для образования водородной связи. Следует отметить, что химические исследования не могли выявить участия Asp-102 в функционировании активного центра, поскольку этот остаток погружен внутрь молекулы. [20]
Независимо от того, является ли алифатическая цепь нормальной или разветвленной, эффективность обратимого связывания алканола ROH на активном центре определяется валовой гидро-фобностью группы R. Все это позволяет рассматривать гидрофобную область активного центра химотрипсина как каплю органического растворителя, расположенную в поверхностном слое белковой глобулы. Эта капля либо адсорбирует гидрофобный ингибитор из воды на поверхность раздела фаз, либо, будучи расположенной несколько углубленно, полностью экстрагирует его. [21]
Эти результаты ( см. рис. 42) интересно сопоставить с данными по структуре кристаллического химотрипсина. Согласно Стейтцу и др. [66], гидрофобная полость в кристаллическом ферменте ( см. рис. 9) действительно узка ( 3 5 - 4 А) и длина ее не превышает 10 А. Таким образом, рентгеновские исследования структуры кристаллического фермента [66] и кинетические исследования его субстратной специфичности в водном растворе [21] привели к взаимно согласующимся данным о геометрии активного центра химотрипсина. [22]
В сущности эти наблюдения и составляют экспериментальную базу доводов Ламри. Но ошибочность этих аргументов становится очевидной, если сравнить актпвационные параметры реакций гидролиза бензоилгирозинамида и бензамида, катализируемого хнмотрипснном и ионом ОН соответственно. Как будет показано ниже, катализ химотрипсином ближе к основному катализу, чем к кислотному или к катализу ионом ОН, по сравнению с Н3О В действительности в данном случае правомерным является любое сравнение, поскольку АЯ и AS очень сильно зависят, например, от природы реагирующих веществ, знака их заряда и, по всей вероятности. ОН - не являются функциональными грунпами, входящими в активный центр химотрипсина. Единственным надежным способом оценки значения эффекта сближения групп в ферментативном процессе было бы сравнение величин ДЯ и AS - для реакции данного субстрата, катализируемой химотрипсином, с теми же величинами бимолекулярной реакции изолированного активного центра химотрипсина с этим же субстратом. [23]
Таким образом, происходящая реакция сводится к образованию промежуточного ацилфермента, а затем к переносу ациль-ного остатка к молекуле воды, если протекает гидролиз, или к иному акцептору ацетильной группы. Серии, конечно, содержится в активном центре не всех ферментов. Так, например, у ри-бонуклеазы его нет, и в этом ферменте, как предполагается, два остатка гистидина выполняют функцию акцептора и донора протонов. Серии и гистидин, несомненно, являются не единственными остатками, необходимыми для работы активного центра. Имеются факты, говорящие об участии триптофана в активных центрах химотрипсина, трипсина, лизоцима, однако об этом пока еще известно мало. [24]
Такие же эффекты наблюдаются при химической модификации фермента низкомолекулярными реагентами. Так, например, ацетилирование и сукцинилирование химотрипсина приводят к смещению оптимума зависимости активности от рН в направлении более щелочных величин рН по сравнению с нативным хи-мотрипсином. Однако это смещение наблюдается только при низкой ионной силе раствора и может быть устранено повышением ионной силы. Это явление, возможно, объясняется возрастанием суммарного отрицательного заряда белка в результате блокирования аминогрупп и соответствующим увеличением рКа имидазоль-ной группы гистидина в активном центре химотрипсина. В случае полиэлектролитного носителя эффекты химической модификации возникают в результате значительно более сильных электростатических эффектов заряженного носителя. [25]
В сущности эти наблюдения и составляют экспериментальную базу доводов Ламри. Но ошибочность этих аргументов становится очевидной, если сравнить актпвационные параметры реакций гидролиза бензоилгирозинамида и бензамида, катализируемого хнмотрипснном и ионом ОН соответственно. Как будет показано ниже, катализ химотрипсином ближе к основному катализу, чем к кислотному или к катализу ионом ОН, по сравнению с Н3О В действительности в данном случае правомерным является любое сравнение, поскольку АЯ и AS очень сильно зависят, например, от природы реагирующих веществ, знака их заряда и, по всей вероятности. ОН - не являются функциональными грунпами, входящими в активный центр химотрипсина. Единственным надежным способом оценки значения эффекта сближения групп в ферментативном процессе было бы сравнение величин ДЯ и AS - для реакции данного субстрата, катализируемой химотрипсином, с теми же величинами бимолекулярной реакции изолированного активного центра химотрипсина с этим же субстратом. [26]
Хотя активный центр относительно невелик, он должен все же представлять собой довольно сложную структуру. Известно, что он определяет и каталитическую активность, и специфичность, а поэтому должен обеспечить весьма тесное взаимодействие, точное в пространственном ( геометрическом) и химическом отношении с молекулами субстрата или с их необходимыми частями. Для проявления активности этого центра необходима его трехмерная структура, кооперативное действие его различных участков, возникающее при их топографическом сближении и соответствующей ориентации. Следовательно, необходима определенная трехмерная структура всей молекулы фермента. В настоящее время принято считать, что активный центр не располагается в пределах какого-либо небольшого отрезка одной пептидной цепи, а представляет совокупность групп, расположенных на двух или нескольких цепях или на различных участках одной, но сложно изогнутой пептидной цепи. Нейратом ( рис, 12), На модели черными линиями показан активный центр химотрипсина, который занимает небольшую область и включает два остатка гистидина и один остаток серина. Здесь имеется одна единственная пептидная цепь, изогнутая таким образом, что различные участки ее ( различные аминокислотные остатки) сближены и образуют каталитически активный центр. Ясно, что каталитическая способность химотрипсина зависит не только от наличия тех или иных функциональных групп, но главным образом от конфигурации всей макроструктуры белка, поскольку эта конфигурация определяет взаимное расположение групп активного центра. Отсюда ясно и значение стабильности макроструктуры ( третичной структуры) белка для выявления и сохранения ферментативной активности. [27]
Страницы: 1 2