Комплементарные цепи
Cтраница 2

Структурные формулы ряда тРНК сейчас уже известны. На схеме обе комплементарные цепи, каждая длиной в 77 звеньев, разбиты па блоки длиной от 8 до 12 нуклеотидов. Эти блоки были синтезированы по заданной программе методами класспч. Далео соседние блоки соединялись друг с другом п стыковка их производилась ферментативным путем с помощью иолинуклеотидлигазы. [17]
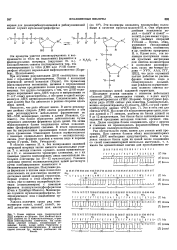
Структурные формулы ряда тРНК сейчас уже известны. На схеме обе комплементарные цепи, каждая длиной в 77 звеньев, разбиты на блоки длиной от 8 до 12 нуклеотидов. Эти блоки были синтезированы по заданной программе методами классич. Далее соседние блоки соединялись друг с другом и стыковка их производилась ферментативным путем с помощью полинуклеотидлигазы. [19]
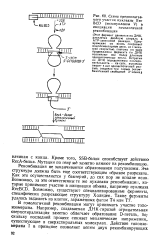 |
Схема предполагаемого участия нуклеазы Rec BCD ( экзонуклеазы V в инициации гомологичной рекомбинации. [20] |
Этот фермент движется по ДНК, расплетая двойную спираль в АТР-завнсимой реакции. Одновременно он вновь спаривает комплементарные цепи, но с меньшей скоростью, поэтому по мере продвижения фермента по ДНК возникают все более длинные одноцепочечные петли. [21]
Длина синтезированных in vitro молекул обычно составляет 20 - 30 нуклеотидов и редко превышает 100 нук-леотидов. Для получения двухцепочечных молекул комплементарные цепи синтезируют по отдельности и затем проводят отжиг. Полученную таким способом ДНК используют в качестве зондов для скрининга геномных библиотек; в качестве линкеров и адаптеров при клонировании генов; для мутагенеза in vitro; для конструирования генов с целью последующего клонирования. [22]
 |
Схема предполагаемого участия нуклеазы Rec BCD ( экзонуклеазы V з инициации гомологичной рекомбинации. [23] |
Этот фермент движется по ДНК. Одновременно он вновь спаривает комплементарные цепи, но с меньшей скоростью, поэтому по мере продвижения фермента по ДНК возникают все более длинные одноцепочечные петли. [24]
ДНК ко-валентно присоединены вирус-специфические белки. Весьма своеобразны концы генома вируса осповакцины и других поксвирусов, а также некоторых иридовирусов - комплементарные цепи ДНК ковалентно соединены меэюду собой, причем как на самом конце молекулы, так и на непосредственно прилегающих к нему участках имеется несколько неспаренных ( некомплементарных) нуклеоти-дов; кроме того, имеется инвертированный концевой повтор значительной длины. Происхождение и значение всех этих особенностей концевых структур станут понятными после рассмотрения способов репликации геномов соответствующих вирусов. [25]
ДНК ко-валентно присоединены вирус-специфические белки. Весьма своеобразны концы генома вируса осповакцины и других поксвирусов, а также некоторых иридовирусов - комплементарные цепи ДНК ковалентно соединены между собой, причем как на самом конце молекулы, так и на непосредственно прилегающих к нему участках имеется несколько неспаренных ( некомплементарных) нуклеоти-дов; кроме того, имеется инвертированный концевой повтор значительной длины. Происхождение и значение всех этих особенностей концевых структур станут понятными после рассмотрения способов репликации геномов соответствующих вирусов. [26]
Механизм разъединения и удвоения отдельных цепей как будто бы совсем прост: ведь цепи удерживаются вместе только при помощи водородных мостиков, которые довольно легко расщепляются, так как энергия такой связи относительно мала. Правда, пока еще точно не известно, откуда исходит приказ расстегнуть молнию и начать строить комплементарные цепи. Достоверно известно только одно: приказ должен быть выполнен до того, как начнется деление или почкование клеток. [27]
Для того чтобы приблизить ее к природе, будем считать, что макромолекулы не автокоппруются, но синтезируют комплементарные цепи. [28]
Далее следует перестройка одного из концов этой молекулы с образованием заячьих ушей. В таком случае синтез новой цепи будет сопровождаться вытеснением исходной родительской цепи из дуплекса; другим продуктом реакции будет двухнитевая репликативная форма, у которой комплементарные цепи опять ковалентно связаны между собой, но на этот раз уже на другом конце молекулы. Затем происходит регенерация и инверсия З - конца комплементарной цепи. [29]
Вполне вероятно, что явление биологической специфичности вообще есть результат полной и тонкой комплементарное между структурами взаимодействующих молекул. Структуры ферментов компле - ментарны напряженным конфигурациям реагирующих молекул. Гены представляют собой комплементарные цепи нуклеиновых кислот ( разд. [30]
Страницы: 1 2 3